Evolução humana
| Parte de uma série sobre |
| Evolução |
|---|
 Diagrama da divergência dos grupos taxonómicos modernos em relação aos seus ancestrais comuns. |
|
Tópicos fundamentais |
Evolução humana é nome que se dá ao processo de origem e evolução de espécies pertencentes ao gênero Homo, como um clado particular dos hominídeos.[1] O estudo da evolução humana engloba muitas disciplinas científicas, incluindo a antropologia física, primatologia, arqueologia, paleontologia, linguística e genética.[2]
O termo "humano" no contexto da evolução humana, refere-se ao gênero Homo, mas os estudos da evolução humana usualmente incluem outros hominídeos, como os australopitecos. O gênero Homo se afastou dos Australopitecos entre 2,3 e 2,4 milhões de anos na África.[3][4] Os cientistas estimam que os seres humanos ramificaram-se de seu ancestral comum com os chimpanzés - o único outro hominídeo vivo - entre há 5 e 7 milhões de anos. Diversas espécies de Homo evoluíram e agora estão extintas. Estas incluem o Homo erectus, que habitou a Ásia, e o Homo neanderthalensis, que habitou a Europa. O Homo sapiens arcaico evoluiu entre há 400 000 e 250 000 anos.
A opinião dominante entre os cientistas sobre a origem dos humanos anatomicamente modernos é a "hipótese da origem única",[5][6][7] que argumenta que o Homo sapiens surgiu na África e migrou para fora do continente em torno de há 50 000 a 100 000 anos, substituindo as populações de Homo erectus na Ásia e de Homo neanderthalensis na Europa. Hoje é mantida essa ideia, mas, apenas questionada essa origem ser africana e não europeia[8]. Já os cientistas que apoiam a "Hipótese multirregional" argumentam que o Homo sapiens evoluiu em regiões geograficamente separadas.
Atualmente, argumenta-se que uma abordagem científica da evolução humana não deve ser entendida apenas como uma sucessão de processos e efeitos biológicos, favorecendo uma abordagem interdisciplinar que leva em conta aspectos culturais como determinantes fundamentais para a compreensão desse processo. Nesse sentido, a evolução humana é considerada como uma evolução biocultural.[1]
Mudanças anatômicas[editar | editar código-fonte]
A evolução humana desde a sua separação inicial do último ancestral comum entre humanos e chimpanzés é caracterizada por várias alterações morfológicas, de desenvolvimento, fisiológicas e comportamentais. Destas, as adaptações mais significativas são o bipedalismo, aumento do tamanho do cérebro, a ontogenia prolongada (gestação e infância) e a redução do dimorfismo sexual. A relação entre essas mudanças é objeto de debate constante. Outras alterações morfológicas significativas incluem a evolução da ação de segurar e agarrar com firmeza (força e precisão), uma mudança que ocorreu primeiramente no H. erectus.[9]
Bipedalismo[editar | editar código-fonte]
Definição[editar | editar código-fonte]
A bipedia obrigatória consiste em se locomover exclusivamente sobre os dois membros inferiores, mantendo a coluna ereta e as mãos livres, é a principal característica que define a tribo Hominini a qual inclui a espécie Homo sapiens e seus ancestrais. A bipedia implicou em uma série de adaptações anatômicas, musculares e ósseas que permitiram a nós este tipo de locomoção.[10] Humanos atualmente movimentam os membros inferiores que estão relativamente duros de maneira que o centro de gravidade que no ponto mais baixo que é o calcanhar, ao ser erguido para o maior ponto em distância média do movimento ao qual tem intenção, gere um movimento estilo pêndulo invertido que permite uma mudança efetiva de potencial gravitacional e energia cinética. O mesmo estilo de caminhar foi adotado por outros bípedes muito antes.[11] As diferentes posturas, levam a diferentes ângulos de quadril, joelhos e tornozelos. O quadrupedalismo é a base para o bipedismo, conforme pressões ambientais foram selecionando novas posturas, foi assim surgindo esse comportamento.[11]
Hipóteses para a origem do bipedalismo[editar | editar código-fonte]
- Hipótese da savana: sendo a principal teoria sobre a origem da bipedia, leva em consideração que uma postura ereta facilitaria a visualização de predadores e presas, diminuiria a área do corpo exposta ao sol evitando hipertermia e pouparia energia no deslocamento visto que o andar sobre duas pernas é menos custoso que correr sobre quatro. esta se popularizou com a descoberta do Australopithecus africanus por Raymond Dart (1925).[12] Segundo esta hipótese, a expansão de savanas foi um fator importante para a evolução hominínia.[13] Ancestrais humanos deixaram as florestas e passaram a ocupar ambientes abertos com algumas regiões florestais e baixa disponibilidade de água [12] - savanas - onde se adaptaram e desenvolveram a postura ereta e o bipedalismo. Outras hipóteses foram integradas à hipótese da savana, pois também buscavam explicar a origem do bipedalismo neste mesmo cenário.[14] O registro fóssil mais antigo de um hominínio com locomoção sobre duas pernas é datado de 7 milhões de anos, o Sahelanthropus tchadensis.[10] Os não bípedes que caminham usando as articulações dos dedos das mãos como apoio - gorilas e chimpanzés - divergiram da linha hominínia neste mesmo período, o que faz do Sahelanthropus ou do Orrorin prováveis candidatos a último ancestral comum. O Ardipithecus, completamente bípede, surgiu há aproximadamente 5,6 milhões de anos.[15] Em 2001 foi encontrado o fóssil do Sahelanthropus tchadensis e, embora o achado se limitasse ao crânio e dentes, foi possível afirmar, a partir da posição do forame magno, que se tratava do primeiro hominídeo com locomoção sobre duas pernas. Um ano antes desta descoberta, foi descoberto no Quênia o fóssil do Orrorin tugenensis (datado de 6 milhões de anos), eram dentes e dois ossos do fêmur cuja curvatura indicava que aquele indivíduo era bípede.[16] Esses achados mostraram que a forma de locomoção sobre duas pernas é anterior ao surgimento de gorilas, chimpanzés e humanos. A filogenia molecular sugere que por volta de 6,4 ± 1,5 milhões de anos a linhagem de gorilas divergiu da linhagem de chimpanzés e humanos, e que humanos e chimpanzés começaram a divergir por volta de 5.4 ± 1,1 milhões de anos.[17] Registros fósseis de Ardipithecus ramidus são datados de 5.8 a 5.4 milhões de anos, coincidindo com o período em que ocorreu a divergência entre chimpanzés e humanos. Muitos estudos sobre a anatomia Ardipithecus, mostram que se tratava de um hominíneo com características tanto de bipedalismo quanto de arborícola corroborando para o estabelecimento deste como o último ancestral comum entre chimpanzés e humanos. A evolução dividiu esta linhagem em duas: de um lado os primeiros hominínios, os quais tornaram-se bípedes obrigatórios e de outro o chimpanzé e seus ancestrais, que se tornaram grandes escaladores vivendo majoritariamente nas árvores.
- Teoria da liberação das mãos: muitas teorias se acumulam sobre o que teria levado ao surgimento e evolução de uma postura ereta e modo de locomoção sobre duas pernas. Um dos modelos teóricos sobre a origem do bipedalismo foi o modelo de Charles Darwin o qual afirma em seu livro "The Descent of Man", and "Selection in Relation to Sex", que: “as mãos e os braços dificilmente poderiam ter se tornado perfeitos o suficiente para ter armas fabricadas, ou para ter atirado pedras e lanças com uma pontaria verdadeira, enquanto fossem habitualmente usados para locomoção e para suportar todo o peso do corpo, ou, como antes comentados, enquanto fossem especialmente adaptados para subir em árvores.”.[18] A afirmação de Darwin implica que a bipedia liberaria as mãos para manuseio de armas e ferramentas associando este fato a diminuição dos dentes caninos. Contudo, a teoria de Darwin sobre o bipedalismo não se sustenta, uma vez que os registros fósseis demostram que o uso de ferramentas e a diminuição dos caninos ocorreram muito tempo após o surgimento da bipedalismo.[14] Baseado na teoria de Darwin, Owen Lovejoy propõe uma teoria de liberação das mãos relacionada a monogamia e a coleta de alimentos. Ele argumenta que andar sobre duas pernas liberaria as mãos para transportar os alimentos com maior eficiência e argumenta também que as fêmeas se ocupariam de cuidar da prole e os machos eram responsáveis por prover as fêmeas com os alimentos coletados. Em contrapartida, as fêmeas se acasalariam somente com estes machos provedores, melhorando a taxa de sobrevivência da prole. Assim os machos não lutariam mais com outros machos e assim os caninos diminuiriam com o tempo. A teoria de Lovejoy é apoiada por evidências antropológicas que usam chimpanzés como modelos de comportamento.[14]
- Teoria da eficiência energética: correlaciona o bipedalismo a baixos custos energéticos. Nesse sentido, Kramer (2009) afirma que o bipedalismo diminuiria a energia dispendida na locomoção em busca de recursos alimentares e aumentaria a disponibilidade de energia para outras funções como a reprodução. Assim, a seleção natural atuaria de modo a diminuir cada vez mais os custos de locomoção. Ainda segundo este autor, a redução de energia poderia ter duas formas: uma seria a eficiência energética (mais atividades realizadas com menos energia) e outra seria a economia de energia (quantidade absoluta de energia utilizada para realizar a atividade).[19] Um estudo comparativo entre humanos e chimpanzés afirma que características anatômicas, como um quadril mais largo e membros inferiores mais longos, teriam proporcionado a maior extensão dos membros inferiores dando mais eficiência energética na caminhada do ancestral comum de chimpanzé e humanos.[20] A hipótese de Sockol pressupõe que o tamanho do membro influi na eficiência energética envolvida na locomoção contudo, outro estudo que explora o bipedalismo em australopitecíneos, afirma que o tamanho dos membros inferiores (semelhante ao tamanho dos membros inferiores de chimpanzés e possivelmente do ancestral comum chimpanzés e humanos) não implicam em maior ou menor eficiência de energia para locomoção, nem afetam a bipedia ou a eficiência na forma de caminhar destes hominídeos, apenas reduz sua velocidade de caminhada.[19] A busca por recursos alimentares, geralmente dispersos no ambiente (seja caçando ou coletando), impunha aos primeiros hominídeos percorrer uma dada distância durante o dia, assim, a velocidade da locomoção tornava-se um fator importante na evolução dos caracteres locomotores. Portanto, a seleção atuaria para tornar um bipede tão veloz quanto deveria ser para percorrer as distâncias necessárias para conseguir recursos e, ao mesmo tempo, reduzia a quantidade de energia necessária para esta atividade.[21] A comparação entre hominídeos mostrou que a eficiência energética e a economia de energia está muito relacionada a forma e tamanho, mas esta relação não é totalmente conhecida, uma vez que o gasto energético varia conforme a espécie, dentro da mesma espécie e ao longo do tempo.[21]
- Hipótese da locomoção em corpos d’água rasos ou da vadeação (em inglês: Wading hypothesis): foi inicialmente proposta em 1960, por Alister Hardy.[22] Ancestrais humanos teriam habitado margens de corpos d’água e adentravam neles em busca de alimento. Deste modo, propõe Hardy: “Parece provável que o Homem aprendeu a ficar em posição ereta inicialmente na água e, à medida que seu equilíbrio melhorou, ele descobriu que estava mais bem equipado para permanecer em pé nas margens quando saía da água e também para correr” (tradução dos autores, p. 644).[22] Influenciada por Hardy, surge a hipótese do macaco aquático segundo a qual a linhagem hominínia passou a habitar temporariamente um ambiente semi-aquático, o que poderia ser a origem, dentre outras adaptações, do bipedalismo.[17] Esta proposta, porém, foi criticada por suas inconsistências com o registro fóssil e por buscar evidências na anatomia do humano moderno.[23]
- Hipóteses integradas à hipótese da savana:
- Hipóteses de display: integrada à hipótese da savana, considera que a postura bípede é adotada a fim de tornar o indivíduo o maior possível.[14] Funciona como uma estratégia de defesa contra predadores, mas também é uma maneira de resolver conflitos dentro do grupo de maneira ritualística e gestual, sem a necessidade de ataques físicos.[17]
- Hipótese do comportamento de sentinela: foi inicialmente proposta por Dart em 1959.[17] Segundo esta hipótese, a postura bípede consiste em uma vantagem evolutiva pois permite que um indivíduo observe seus arredores e identifique possíveis predadores. Em um estudo sobre o comportamento de babuínos realizado por Rose (1976), observou-se que o comportamento de alerta é a instância de postura bípede mais frequente, logo após a postura bípede durante a alimentação.[24] Rose adota os babuínos como modelo e propõe que a postura bípede seria a base sobre a qual a seleção natural atuaria. Assim como Rose, Hunt (1994) [25] investiga contextos que evocam bipedalismo a fim de estudar pressões seletivas e observa que padrões comportamentais bípedes associados à alimentação eram os mais comuns em sua pesquisa com chimpanzés. Ele aponta que o comportamento sentinela era raro e não traria vantagens suficientes que compensassem as desvantagens de um bipedalismo inicial e pouco adaptado. Hunt também indica que ainda não foi demonstrado que a postura bípede aumentaria o campo de visão em ambientes de floresta, habitat onde se hipotetiza que a transição para o bipedalismo ocorreu.
- Hipótese da coleta de sementes: proposta por Cliff Jolly (1970),[26] foi elaborada a partir de um modelo com babuínos-gelada. Foi observado que assumiam uma posição bípede em cócoras enquanto se alimentavam, o que liberava as mãos para coletar folhas de grama, sementes e rizomas. Jolly propõe que a adaptação à vida terrestre ocorreu também com a transição de uma dieta baseada em frutas ou folhas para cereais, alimentos ricos em calorias. A manutenção da postura bípede teria facilitado a evolução da locomoção bípede, pois alguns elementos do bipedalismo já comporiam o repertório comportamental. Wrangham (1980) [27] apontou que, apesar da pouca eficiência do caminhar em cócoras de babuínos-gelada, este comportamento ainda é frequente. Um possível benefício é que esta postura permite que continuem a se alimentar com as mãos sem interromper o forrageamento. Também sugere que a manutenção desta postura reduz a necessidade de se abaixar e se levantar enquanto alterna entre postura bípede e quadrúpede, o que pode ter tornado o bipedalismo facultativo mais frequente em função da eficiência energética.
- Hipótese da termorregulação: em 1967, Ward e Underwood [28] realizaram uma pesquisa a respeito do efeito da postura em relação à área de superfície do corpo exposta à radiação solar. Em regiões de baixa latitude, a postura bípede parece ser mais vantajosa. Wheeler, considerando o problema da hipotermia em ambientes de temperaturas elevadas de um cenário de savana, propõe a hipótese de que o bipedalismo foi selecionado por favorecer a termorregulação. Wheeler se apoia nas pesquisas de Ward e Underwood ao afirmar que uma postura bípede diminui a incidência de radiação solar, o que diminui o ganho de calor. A postura bípede também permitiria acesso a um micro-habitat mais fresco,[25] com acesso a ventos e temperaturas mais favoráveis ao se distanciar do chão. A vantagem termorreguladora do bipedalismo permitiria, então, melhor exploração de ambientes quentes e favoreceria sua fixação.
Novas descobertas paleoclimatológicas, porém, apontam que a transição para o bipedalismo ocorreu em ambientes de floresta, bem como fósseis com os primeiros sinais de bipedalismo ainda apresentavam adaptações ao deslocamento arborícola.[13][14][25][29] Sussman e Hart (2008)[29] apontam que, embora o clima do continente africano tenha começado a se tornar mais árido entre 12 e 5 milhões de anos atrás, ainda restavam áreas de transição entre florestas e savana, onde se iniciaram mudanças comportamentais e anatômicas na evolução hominínia. Deste modo, as hipóteses da savana precisam ser repensadas e recontextualizadas dentro da linha do tempo da evolução hominínia.
Diferentemente das propostas anteriores, há a hipótese de que diversos fatores influenciaram o surgimento do bipedalismo.[14][17] Embora muitas propostas busquem encontrar a causa definitiva que levou à adoção de uma postura e deslocamento bípede, esta investigação é dificultada pelo fato de que os efeitos da seleção natural são observados em uma larga escala, dentro da qual diversos fatores podem interferir.[14] Segundo Niemitz, “a pergunta que uma hipótese promissora deve responder é qual combinação das pressões seletivas ultrapassou o ponto a partir do qual a adoção habitual de postura ereta e o deslocamento bípede contínuo passou a valer a pena” (tradução dos autores, p. 250). Se atribuídas ao período adequado na história evolutiva dos hominínios, diversas hipóteses, ou ao menos alguns aspectos, podem contribuir para o entendimento de como o bipedalismo evoluiu para seu modo atual.[17] As hipóteses da savana, por exemplo, poderiam explicar porque o bipedalismo se tornou cada vez mais dominante, a ponto de substituir o bipedalismo parcial hominínio.[14]
Assim, é possível que o bipedalismo tenha sido favorecido porque liberou as mãos para alcançar e transportar alimentos, tornou a locomoção[30] mais eficiente do ponto de vista energético, possibilitou caçadas e corridas de longa distância, forneceu um campo de visão aprimorado, além de ajudar a evitar a hipertermia por reduzir a área de superfície do corpo exposta diretamente ao sol. Todos esses são recursos vantajosos para prosperar no novo ambiente de savana e bosque resultantes da elevação do Vale do Rift na África Oriental ao invés do habitat anterior de floresta fechada.[30][31][32] No entanto, estudos recentes sugerem que a bipedalidade sem a capacidade de usar o fogo não teria permitido a dispersão global.[33]
Adaptações anatômicas[editar | editar código-fonte]
- Ossos pós-cranianos: várias adaptações relacionadas à forma de locomoção de um animal são observadas em seu esqueleto. Em animais bípedes a região inferior do esqueleto (pélvis e membros inferiores) requer estruturas adaptadas para suportar o peso do corpo. Comparativamente aos mamíferos quadrúpedes, a pélvis adaptada à bipedia possui formato de bacia, com ossos ilíacos mais curtos e largos, acomodando melhor os órgãos e mantendo o peso e estabilidade dos movimentos alternados das pernas. Além disso, os joelhos possuem articulações que permitem a extensão total da perna, e o polegar dos pés não ocorre de forma opositora.[10] Harcourt-Smith e Aiello (2004), em um trabalho de revisão sobre fósseis de pés de primatas, argumentam que as origens e a evolução da locomoção bípede podem ser melhor compreendidas quando se estabelecem relações entre a estrutura e função dos ossos dos pés de ancestrais hominídeos e primatas existentes. Isto se deve à especialização destas estruturas e sua importância no desenvolvimento da bipedia, já que, neste modo de locomoção, os pés passam a ser a única estrutura em contato direto com o solo e, consequentemente, sofreram pressões seletivas em relação à eficiência do equilíbrio e da propulsão.[34]
As mudanças mais significativas ocorreram na região pélvica, onde o osso ilíaco voltado para baixo foi encurtado e ampliado como requisito para manter o centro de gravidade estável durante a caminhada;[35] os hominídeos bípedes têm uma pelve mais curta, porém mais larga, em forma de tigela, devido a isso, o que leva a uma desvantagem aos hominídeos não bípedes: o canal de nascimento dos hominídeos bípedes é menor do que nos hominídeos não bípedes, embora tenha havido um alargamento se compararmos os australopitecíneos com os humanos modernos para permitir a passagem de recém-nascidos devido ao aumento do tamanho do crânio, mas isso é limitado à porção superior, pois um aumento adicional poderia dificultar o movimento bipedal normal.[36]
O encurtamento da pelve e o canal de nascimento menor evoluíram como requisitos para o bipedalismo e tiveram efeitos significativos no processo de nascimento humano, que é muito mais difícil nos humanos modernos do que em outros primatas. Durante o nascimento humano, devido à variação no tamanho da região pélvica, a cabeça do feto deve estar em uma posição transversal (em comparação com a mãe) durante a entrada no canal do parto e girar cerca de 90 graus após a saída.[37] O canal de nascimento menor tornou-se um fator limitante para o aumento do tamanho do cérebro nos primeiros humanos e levou a um período de gestação mais curto, levando à imaturidade relativa da prole humana, que não consegue andar muito antes dos 12 meses e têm maior neotenia, comparado com outros primatas, que são ágeis numa idade muito menor.[32] O aumento do crescimento cerebral após o nascimento e o aumento da dependência das crianças em relação às mães tiveram um efeito importante no ciclo reprodutivo feminino,[38] e o surgimento mais frequente de aloparentalidade em humanos quando comparado com outros hominídeos.[39] O atraso na maturidade sexual humana também levou à evolução da menopausa, o que sugere a explicação de que as mulheres idosas poderiam transmitir melhor seus genes cuidando da prole da filha, ao invés de ter mais filhos.[40]
- Ossos cranianos: em relação a estrutura craniana, a bipedia pode ser inferida através do posicionamento centralizado do forame magno, onde se conecta a coluna vertebral.[10] No entanto, alguns fósseis necessitam de informações do esqueleto pós-craniano para a afirmação de bipedia. Como exemplo, em um trabalho de reconstrução virtual craniana de Sahelanthropus tchadensis, Zollikofer e colaboradores (2005)[41] observaram a relação angular entre o foramen magno e o plano orbital desta espécie e de outras três (Homo sapiens; Pan troglodytes e Australopithecus africanus), mas concluíram que mais informações são necessárias para afirmar que se trata de uma espécie bípede. Em 1924, um fóssil de Australopithecus africanus (conhecido como "a criança de Taung") foi descrito como o primeiro australopitecínio e considerado um hominínio a partir de sua anatomia craniana. Quatorze anos depois, um fóssil de Paranthropus robustus com características cranianas indicativas de bipedia, foi considerado parte de uma linhagem importante na evolução dos hominíos.[10]
Encefalização[editar | editar código-fonte]

A espécie humana acabou desenvolvendo um cérebro muito maior que o de outros primatas - tipicamente 1 330 cm³ nos humanos modernos, quase três vezes o tamanho do cérebro de um chimpanzé ou gorila.[42] Após um período de estase no Australopithecus anamensis e Ardipithecus, espécies que tinham cérebros menores resultantes de sua locomoção bípede,[43] o padrão de encefalização começou com o Homo habilis, cujo cérebro de 600 cm³ era um pouco maior que o dos chimpanzés. Esta evolução continuou no Homo erectus com cérebros entre 800 e 1 100 cm³, e atingiu o máximo nos neandertais com cérebros entre 1 200 e 1 900 cm³, maiores até mesmo que o cérebro dos modernos Homo sapiens. Este aumento de volume ocorre durante o crescimento do cérebro após o nascimento e é muito superior ao dos outros macacos (heterocronia). Também permitiu um período mais extenso de aprendizado social e aquisição de linguagem nos humanos jovens, começando há cerca de 2 milhões de anos.
Além disso, as mudanças na estrutura dos cérebros humanos podem ser ainda mais significativas que o aumento no tamanho.[44][45][46][47]

Os lobos temporais, que contêm os centros de processamento da linguagem, cresceram desproporcionalmente, assim como o córtex pré-frontal, que é relacionado à tomadas de decisão complexas e à moderação do comportamento social.[42] Atribui-se à encefalização um aumento na dieta de carnes e amido,[48][49][50] e o desenvolvimento do cozimento dos alimentos.[51] Também tem sido proposto que a inteligência aumentou como uma resposta à necessidade crescente de resolver problemas sociais à medida que a sociedade humana se tornou mais complexa.[52] Alterações na morfologia do crânio, como mandíbulas menores e pontos de fixação dos músculos, forneceram mais espaço para o cérebro crescer.[53]
O aumento no volume do neocórtex também inclui um rápido aumento no tamanho do cerebelo. Sua função tem sido tradicionalmente associada ao equilíbrio e ao controle motor fino, mas mais recentemente com a fala e a cognição. Os grandes macacos, incluindo os hominídeos, têm um cerebelo mais pronunciado em relação ao neocórtex do que os outros primatas. Sugere-se que, devido à sua função de controle sensorial-motor e ao aprendizado de ações musculares complexas, o cerebelo deva ter apoiado as adaptações tecnológicas humanas, inclusive as pré-condições para a fala.[54][55][56][57] A este respeito, é importante notar que a Neurobiologia é uma entre várias frentes de investigação que se dedicam ao estudo da origem da linguagem humana. Para compreender de maneira científica o fenômeno da linguagem em humanos e sua evolução, é preciso articular evidências e hipóteses ligadas a áreas que variam da Linguística à Paleoantropologia, da Etologia à Genética, entre muitas outras.[58]
A vantagem imediata de sobrevivência da encefalização é difícil de discernir, pois as principais alterações cerebrais do Homo erectus para o Homo heidelbergensis não foram acompanhadas por grandes mudanças na tecnologia. Sugere-se que as alterações foram principalmente sociais e comportamentais, incluindo o aumento das habilidades empáticas,[59][60] do tamanho dos grupos sociais[61][62][63] e da plasticidade comportamental.[64] A encefalização pode também dever-se a uma dependência de alimentos ricos em calorias e difíceis de adquirir.[65]
Dimorfismo sexual[editar | editar código-fonte]
Dimorfismo é definido como “a existência de dois ou mais tipos distintos de morfo-fisiologias dentro de uma mesma espécie”, nesse sentido, o dimorfismo sexual diz respeito às diferenças biológicas entre fêmeas e machos, abrangendo características diversas, não somente ligadas ao sistema reprodutivo em si.[66]
A espécie humana, quando comparada a outras espécies de primatas não humanos, apresenta diferenças entre os sexos mais moderadas.[67] Por exemplo, exibimos diferenças pequenas na estatura, no peso, e principalmente há grande redução dos caninos nos homens tornando-os semelhantes aos das mulheres, característica que claramente nos diferencia dos outros primatas.[68]
Em se tratando da reprodução e das práticas sexuais humanas, também há muita distinção comparando-as às dos outros primatas. O processo evolutivo promove a manutenção das espécies, o que depende da capacidade de deixar descendentes. A principal força evolutiva envolvida neste caso é a seleção sexual, que atua favorecendo características que aumentam o sucesso reprodutivo, sendo, então, um mecanismo que possibilita que indivíduos transmitam seus genes. Ela é responsável, também, por moldar a fisiologia de fêmeas e machos, e, como Darwin observou, ela pode ser transformada, moldada e limitada por práticas socioculturais. Em geral, quando se trata de humanos, além desse aspecto, outros parâmetros devem ser levados em consideração: biológicos, ambientais, materiais e comportamentais.[69]
O comportamento humano ligado às relações sexuais e de parceria (como o casamento) pode ter sido um dos maiores fatores que moldou a seleção dos caracteres sexuais secundários (as característica sexualmente dimórficas), mas o contrário também pode ocorrer.[69] As diferenças entre os sexos podem ser atribuídas, então, a diferenças acentuadas na estratégia reprodutiva de cada um.[70] Há, contudo, um padrão universal destas características,[70] o que sugere uma seleção muito antiga no tempo evolutivo.
Uma característica que influenciou grandemente a maneira como os humanos se relacionam é a ovulação oculta: nas fêmeas humanas não há quase que nenhum sinal que indica o momento de receptividade (o momento da ovulação), não há manifestações internas (que ocorrem em outros primatas não humanos) da sua mudança hormonal. Essa fisiologia nos torna capazes de ter relações sexuais a qualquer momento, independente do período fértil.[69] Pesquisas sociológicas indicam que isso pode ter sido um recurso que reforçou o compromisso dos pais dentro das famílias, forjando laços de lealdade paterna, pois seria mais difícil confirmar a paternidade da prole sem ter certeza se houve cópula entre marido e mulher durante a ovulação.[69]
Entre humanos existem diversos fatores que podem influenciar o dimorfismo sexual, como “peso, altura, pelagem, rosto, musculatura, voz, forma corporal, tamanho dos olhos e comportamento”.[71]

Há uma grande diferença de tamanho entre machos e fêmeas humanos, ilustrada pela estatura. Essa diferença, onde os homens são em média 7% mais altos que as mulheres, pode ser atribuída também às relações de proporção do corpo. A deposição de gordura e musculatura diferencial afeta a composição corporal do corpo adulto, acarretando em diferenças visíveis onde mulheres tendem a uma forma corporal “violão” e a dos homens seria semelhante a um triângulo invertido, o que também está ligado a distinção entre as formas dos ossos, principalmente da pélvis.[70] A distinção hormonal durante a adolescência é um importante fator que gera esses padrões de crescimento divergentes, uma vez que meninos e meninas acumulam diferentes quantidades de massa magra e gordura durante esse período.[72] Nessa época do desenvolvimento, as mudanças na composição corporal resultam em homens apresentando mais área muscular e mulheres apresentando um aumento do acúmulo de gordura. Essas diferenças são apontadas como distribuições de tecido adiposo andróide (parte superior do corpo, tronco) versus ginóide (quadril e coxa). Assim, ao atingir a idade adulta homens e mulheres possuem um dimorfismo considerável em sua composição corporal total, com mulheres tendo uma porcentagem média maior de gordura enquanto homens possuem mais massa magra, sendo tal relação observada em todas as populações de grupos étnicos.[70] No entanto, esse dimorfismo não permanece estável na idade adulta, pois com o aumento da idade ocorre perda de massa muscular devido a mudanças hormonais, menor nível de atividade e maior acúmulo de gordura.[70][73]
O rosto humano também desempenha um papel significativo no aspecto do dimorfismo sexual, em Samal et al. foi demonstrado que cerca de 85% das características faciais são bastante divergentes entre machos e fêmeas. Em estudos anteriores, como o de Ferrario et al., foi observado que na maioria dos casos o rosto feminino é menor quando comparado ao masculino, e que as partes média e inferior do rosto mostram mais variação de gênero. O dimorfismo sexual da face humana não é significativo em crianças pequenas, mas se torna cada vez mais significativo à medida que se aproximam da idade adulta, ou seja, muda ao longo do tempo.[10]
Uma outra característica muito perceptível seria a diferença o tom de voz, em que os homens apresentam tons mais baixos, enquanto que as mulheres conseguem alcançar notas mais altas.[74] Embora as causas anatômicas para a diferença do tom de voz em machos sejam conhecidas, não há um consenso acerca da evolução dessa característica. A visão mais tradicional é de que um tom de voz mais baixo seria vantajoso na competição intrassexual, pois existiria uma percepção na fêmea entre uma voz mais grave e a dominância social junto com o porte do corpo do macho.[75] Todavia, estudos empíricos mais recentes têm indicado que a relação entre tamanho de corpo e voz pode ser ecológicamente irrelevante.[76] Além disso, outra grande característica é a distribuição distinta de pelos corporais.[67]
O desenvolvimento diferenciado dessas características sexualmente dimórficas é regido pelos hormônios sexuais. Durante a puberdade, hormônios andrógenos adrenais levam a uma aceleração do crescimento em altura, da maturação óssea e levam também à distribuição de gordura, todas essas características ocorrem em ritmos diferentes, e em regiões do corpo diferentes em garotos e garotas. Os hormônios testosterona e o estrogênio também auxiliam na deposição de gordura localizada, nas regiões abdominal e glúteo-femoral, respectivamente. A testosterona especificamente age sobre o aumento de massa magra que ocorre durante a puberdade, facilitando garotos dessa idade a formar musculatura mais facilmente.[70] Já o estrogênio está envolvido na formação da massa óssea, e estudos têm reconhecido o envolvimento deste na formação de gordura: o tecido adiposo possui receptores para estrógeno, mas a expressão destes é diferente entre os depósitos de gordura visceral e subcutâneos.[77]
Oposição ulnar[editar | editar código-fonte]
A oposição ulnar - o contato entre o polegar e a ponta do dedo mindinho da mesma mão - é único ao gênero Homo,[78] incluindo os neandertais, os hominínios de Sima de los Huesos e os humanos anatomicamente modernos.[79][80] Em outros primatas, o polegar é curto e incapaz de tocar a ponta do dedo mindinho.[79] A oposição ulnar facilita o agarrar com precisão e força da mão humana, subjacentes a todas as habilidades de manipulação.
Outras mudanças[editar | editar código-fonte]
Várias outras mudanças também caracterizaram a evolução dos seres humanos, entre elas uma maior importância na visão, em vez de no olfato; um período de desenvolvimento juvenil mais longo e maior dependência infantil; um intestino menor; o metabolismo basal mais rápido;[81] perda de pelos do corpo; evolução das glândulas sudoríparas; uma mudança na forma da arcada dentária em formato de U para um formato parabólico; desenvolvimento de um queixo (encontrado apenas no Homo sapiens); desenvolvimento de processos estiloides; e o desenvolvimento de uma laringe descendente.
Evolução da linguagem[editar | editar código-fonte]
Precursores da linguagem em primatas[editar | editar código-fonte]
A origem da linguagem é multifatorial e em primatas convém distinguir linguagem de vocalização. Linguagens são construídas a partir de um repertório de sons aprendidos e modificados, que se combinam em sílabas, que se combinam em palavras, que se combinam em frases. O sentido das palavras em uma frase deriva não só do sentido da palavra em si, mas também da palavra associada com as demais palavras da frase. Então, a partir de regras gramaticais, é possível que um número limitado de elementos transmita um número quase infinito de significados. Vocalização, entretanto, possui um número muito mais reduzido de sons e não há muitas evidências que indiquem uma articulação entre esses sons. Assim, se comparada a linguagens, acredita-se que a vocalização de primatas transmita um conjunto de informações mais limitado.[82]
Primatas, humanos e não-humanos, compartilham mecanismos neurológicos para percepção, processamento e resposta à comunicação, como reconhecer faces, vozes e um processamento integrado entre esses dois fatores. Além disso, considera-se a linguagem como sendo presente dentro de uma circunstância social, assim tendo influências de hierarquia e de contexto. Já existem evidências de comunicação similar a uma forma de linguagem em babuínos, por exemplo, levando em conta a hierarquia e a dinâmica social em grupos desses animais. Essa forma de comunicação apresenta alguns mecanismos cognitivos que se entende como sendo marcadores de separação entre linguagem e comunicação em primatas não-humanos.[82]
Neuroanatomia comparada[editar | editar código-fonte]
O desenvolvimento do léxico é essencial para a aquisição de linguagem. O léxico permite referir-se a elementos externos. Chimpanzés e gorilas, quando treinados desde cedo, podem aprender aspectos da linguagem de sinais, adquirindo um léxico razoável, apesar de não demonstrarem domínio gramatical.[83] Estudos sugerem que a evolução da linguagem em humanos pode estar associada a mudanças na conectividade e no volume de substância branca subjacente à área de Broca.[84] O córtex frontal granular desempenha um papel crucial na organização e controle de funções cognitivas superiores, como a linguagem. Regiões, incluindo a área de Wernicke e áreas inferoparietais conectadas à área de Broca, formam um circuito de memória de trabalho para processar e aprender vocalizações complexas, contribuem para o desenvolvimento da linguagem.[85]
Hipótese do papel da percepção categórica na evolução da linguagem[editar | editar código-fonte]
Recentemente foi proposto a hipótese de que a percepção categórica pode ter estabelecido as bases para a discrição, uma das características fundamentais da linguagem.[86] De acordo com o linguista Charles Hockett, uma das características da linguagem humana é seu caráter discreto e composicional, em que os fonemas das sílabas podem ser reutilizados para a composição de novas sílabas. [86][87] Esta capacidade de combinar os fonemas para codificar significados contrasta com os sinais de animais não humanos, que são combinações fixas de sons. Além disso, o caráter discreto da linguagem humana também pode ser observado a nível sintático.[86]
A percepção categórica é definida como um processo psicofísico no qual entradas contínuas são percebidas de forma discreta em diferentes modalidades e foi inicialmente identificada em sons da fala humana.[86] Estudos indicam que bebês são capazes de discriminar diferentes sons em todas as línguas e, à medida que crescem, agrupam os sons não contrastantes de sua língua materna.[86] Comparações filogenéticas apontam que a percepção categórica já está presente em outros animais não humanos, como primatas não humanos, rãs, aves e inclusive grilos. Isso indica que a percepção categórica pode ser um caráter preservado que serviu de base para o desenvolvimento da discrição da linguagem. Ademais, a demonstração de que a aprendizagem de categorias e a aprendizagem de palavras estão intimamente relacionadas sugere que a discrição pode ter se originado da percepção categórica.[86]
No que diz respeito às evidências neurobiológicas, as atividades ligadas à percepção categórica ativam áreas corticais e subcorticais, incluindo o córtex auditivo e pré-motor e os gânglios basais. A percepção categóricas do sons para a discriminação fonêmica e não fonêmica em humanos é dada pelo giro temporal superior (GTS) e pelo sulco temporal superior (STS). O GTS é conectado pelo caminho dorsal I ao córtex pré-motor, que é ativado em tarefas de categorização de fonemas. O caminho dorsal I serve como base para o caminho dorsal II, envolvido no processamento sintático complexo durante o desenvolvimento. Isso pode indicar que a discrição depende da percepção categórica dos fonemas, que baseia a percepção categórica das palavras. Já os gânglios basais parecem estar envolvidos com a aprendizagem da categorização de fala em adultos (no aprendizado de línguas não nativas).[86]
Combinatoriedade de um sistema vocal[editar | editar código-fonte]
A combinação de unidades vocais básicas para formação de sequências mais complexas também é explorada por pesquisadores ao comparar os sistemas vocais humanos e de espécies de animais não-humanos, principalmente com pássaros, cetáceos e primatas. A ideia de “combinatoriedade” trata da capacidade de criar novas mensagens através da combinação de unidades menores em um sistema de comunicação. Os chamados e sons específicos de cada espécie podem ser utilizados para transmissão de informações que, apesar de diferentes das linguagens humanas mais complexas, possuem propriedades fundamentais de geração de mensagens utilizando unidades básicas que são compreendidas pelo grupo.[88]
O compartilhamento das estruturas neurais e processos cognitivos entre humanos e outros animais não-humanos quanto a combinatoriedade vocal é explorada a partir de um ponto de vista evolutivo, apresentando possíveis conexões entre as capacidades de comunicação linguística entre humanos e outros animais, além de origens da linguagem e vocalização no geral (como na produção de música) humana.[88][89][90] As comparações entre as capacidades vocais e linguísticas de diferentes grupos animais auxiliam na abordagem investigativa do tópico uma que buscar pelas suas origens pode ser dificultado pela falta de traços fósseis da evolução desse caráter.[88][91]
O surgimento de capacidade de combinação vocal em diferentes grupos de animais sugere hipóteses de processos de convergência evolutiva para essa característica a partir de similaridades nas pressões seletivas colocadas sobre diferentes espécies como o favorecimento causado pela comunicação em diversos contextos sociais e ambientais desses animais, incluindo influenciando na sua eficácia de acasalamento, fuga de presas e busca por recursos alimentares. Assim, a combinatoriedade de unidades básicas de vocalização formando mensagens mais complexas poderiam levar a vantagens adaptativas funcionais.[88] Hipóteses a respeito da origem das bases neurais e cognitivas da capacidade de combinatoriedade vocal são associadas às estruturas cerebrais compartilhadas entre humanas e animais não-humanos. Estudos neurobiológicos sugerem a relação entre estruturas neurais envolvidas na compreensão da linguagem humanas com origens do sistema neural mais basal em outros animais, propondo uma continuidade evolutiva.[88]
Aprendizagem Social e Cultura[editar | editar código-fonte]
O processo de aprendizagem pode ser caracterizado como individual, nos casos de “tentativa e erro” e como social, quando ocorre a observação e imitação. No contexto da evolução humana, considera-se que a aprendizagem social apresenta um maior peso, uma vez que o indivíduo se adapta ao ambiente conforme a experiência dos indivíduos ao redor e não somente evolui baseado na sua própria intuição. Com isso, há uma redução do prejuízo adquirido pelo processo de “tentativa e erro", além de promover a evolução cumulativa do comportamento adaptativo pela imitação através de várias gerações, gerando-se a cultura. Aprendizagem social, segundo McGrew e Tutin[92] (1978), requer: inovação, disseminação, estandardização, durabilidade, difusão e tradição. E, de acordo com Tomasello, a transmissão social fiel é importante pois funciona como uma catraca, ou seja, o que é aprendido é aprimorado e evita retrocessos.[93]
|
|
- Evolução humana
-
-
-
Homo habilis, o primeiro a usar ferramentas de pedra.
-
-





Antes do Homo[editar | editar código-fonte]
Evolução inicial dos primatas[editar | editar código-fonte]
A história evolucionária dos primatas remonta a 65 milhões de anos.[94][95][96][97][98] Uma das mais antigas espécies de mamíferos semelhante a primatas, o Plesiadapis, viveu na América do Norte;[99][100][101][102][103][104] outro, o Archicebus, viveu na China.[105] Outros primatas basais semelhantes eram comuns na Eurásia e na África durante os períodos tropicais do Paleoceno e Eoceno.


David R. Begun[106] concluiu que primatas primitivos se desenvolveram na Eurásia e que uma linhagem que leva aos macacos e humanos africanos, incluindo o Dryopithecus, migrou para o sul a partir da Europa ou da Ásia Ocidental para a África. A população tropical sobrevivente de primatas - que é vista mais completamente no Eoceno Superior e nos estratos fósseis mais antigos do Oligoceno da depressão de Faium a sudoeste do Cairo - deu origem a todas as espécies de primatas existentes, incluindo os lêmures de Madagascar, os lóris do sudeste da Ásia, os gálagos da África, e aos antropoides, que são os Platyrrhini ou macacos do Novo Mundo, os Catarrhini ou macacos do Velho Mundo e os grandes símios, incluindo os humanos e os outros hominídeos.
O mais antigo catarrino conhecido é o Kamoyapithecus do final do Oligoceno encontrado em Eragaleit no norte no Vale do Rift, no Quênia, datado de há 24 milhões de anos.[107] Acredita-se que sua ancestralidade esteja ligada a espécies relacionadas com Aegyptopithecus, Propliopithecus e Parapithecus de Faium, de cerca de há 35 milhões de anos.[108] Em 2010, o Saadanius foi descrito como um parente próximo do último ancestral comum do grupo coroa dos catarrinos e provisoriamente datado entre há 29 e 28 milhões de anos, ajudando a preencher uma lacuna de 11 milhões de anos no registro fóssil.[109]

No Mioceno inferior, a cerca de 22 milhões de anos, os muitos tipos de catarrinos primitivos arborícolas da África Oriental sugerem uma longa história de diversificação prévia. Fósseis de 20 milhões de anos incluem fragmentos atribuídos ao Victoriapithecus, o mais antigo macaco do Velho Mundo. Entre os gêneros que se acredita estar levando à linhagem dos grandes macacos há 13 milhões de anos estão Proconsul, Rangwapithecus, Dendropithecus, Limnopithecus, Nacholapithecus, Equatorius, Nyanzapithecus, Afropithecus, Heliopithecus e Kenyapithecus, todos da África Oriental.
A presença de outros não-cercopitecídeos generalizados do Mioceno médio de locais distantes - Otavipithecus de depósitos de cavernas na Namíbia, e Pierolapithecus e Dryopithecus da França, Espanha e Áustria - é evidência de uma ampla diversidade de formas em toda a África e na bacia do Mediterrâneo durante os regimes climáticos relativamente quentes e uniformes do Mioceno inferior e Médio. O mais antigo dos hominoides do Mioceno, o Oreopithecus, foi encontrado em depósitos de carvão na Itália e datado de cerca de há 9 milhões de anos.
Evidências moleculares indicam que a linhagem dos gibões (família Hylobatidae) divergiu da linha de grandes símios há cerca de 18 a 12 milhões de anos, e a dos orangotangos (subfamília Ponginae) divergiu dos outros grandes macacos há cerca de 12 milhões de anos; não há fósseis que documentem claramente a ancestralidade dos gibões, que podem ter se originado em uma população hominoide do sudeste asiático até então desconhecida, mas os proto-orangotangos fósseis podem ser representados pelo Sivapithecus da Índia e pelo Griphopithecus da Turquia, ambos datados de cerca de há 10 milhões de anos.[110]
Divergência do clado humano de outros grandes macacos[editar | editar código-fonte]
Espécies próximas ao último ancestral comum de gorilas, chimpanzés e humanos podem ser representadas pelos fósseis do Nakalipithecus encontrados no Quênia e do Ouranopithecus encontrados na Grécia. Evidências moleculares sugerem que entre há 8 e 4 milhões de anos, primeiro os gorilas, e depois os chimpanzés (gênero Pan) se separaram da linha que levou aos humanos. O DNA humano é aproximadamente 98,4% idêntico ao dos chimpanzés quando comparados polimorfismos de nucleotídeo único. No entanto, o registro fóssil de gorilas e chimpanzés é limitado; a falta de preservação - os solos das florestas tropicais tendem a ser ácidos e dissolvem os ossos - e o viés de amostragem provavelmente contribuem para esse problema.
Outros hominínios provavelmente se adaptaram a ambientes mais secos fora da faixa equatorial e lá encontraram antílopes, hienas, cães, porcos, elefantes, cavalos e outros animais. A faixa equatorial se contraiu depois de cerca de há 8 milhões de anose há escassas evidências fósseis para a divisão - que se acredita ter ocorrido nessa época - da linhagem hominínia das de gorilas e chimpanzés. Os fósseis mais antigos que alguns argumentam pertencer à linhagem humana são o Sahelanthropus tchadensis (cerca de 7 milhões de anos) e Orrorin tugenensis (cerca de 6 milhões de anos), seguidos pelo Ardipithecus (5,5-4,4 milhões de anos), com as espécies Ar. kadabba e Ar. ramidus.
Argumenta-se em um estudo da história de vida do Ar. ramidus que a espécie fornece evidências para um conjunto de adaptações anatômicas e comportamentais em hominínios muito antigos, diferentes de qualquer espécie de grande macaco existente.[111] Este estudo demonstrou afinidades entre a morfologia craniana do Ar. ramidus e de chimpanzés infantis e juvenis, sugerindo que as espécies desenvolveram uma morfologia craniofacial juvenilizada ou pedomórfica via dissociação heterocrônica de trajetórias de crescimento. Argumenta-se também que a espécie apoia a noção de que hominínios muito antigos, da mesma forma que os bonobos (Pan paniscus), a espécies menos agressiva do gênero Pan, podem ter evoluído através do processo de auto-domesticação. Consequentemente, argumentando contra o chamado "modelo referencial do chimpanzé", os autores sugerem que não é mais recomendável usar comportamentos sociais e de acasalamento de chimpanzés (Pan troglodytes) em modelos de evolução social inicial dos hominínios. Ao comentar sobre a ausência de morfologia canina agressiva no Ar. ramidus e as implicações que isso tem para a evolução da psicologia social hominínia, eles escreveram:
É obvio que o Ar. ramidus difere significativamente dos bonobos, porque os bonobos mantiveram caninos funcionais afiados. No entanto, o fato de que o Ar. ramidus compartilha com os bonobos o dimorfismo sexual reduzido e uma forma mais pedomórfica em relação aos chimpanzés, sugere que as adaptações de desenvolvimento e sociais evidentes nos bonobos podem ajudar nas futuras reconstruções da psicologia social e sexual inicial dos hominínios. De fato, a tendência para o aumento do cuidado materno, seleção de parceiras femininas e autodomesticação pode ter sido mais forte e mais refinada no Ar. ramidus do que vemos nos bonobos.[112]
Os autores argumentam que muitas das adaptações humanas básicas evoluíram nos antigos ecossistemas arborizados e florestais do final do Mioceno e do início do Plioceno na África. Consequentemente, eles argumentam que os humanos podem não representar a evolução de um ancestral semelhante ao chimpanzé, como tradicionalmente se supunha. Isso sugere que muitas adaptações humanas modernas representam traços filogeneticamente profundos e que o comportamento e a morfologia dos chimpanzés podem ter evoluído após sua separação do ancestral comum que eles compartilham com os humanos.
Este artigo ou seção pode conter informações desatualizadas. |
Gênero Homo[editar | editar código-fonte]
Na taxonomia moderna, o Homo sapiens é a única espécie existente desse gênero, Homo. Do mesmo modo, o estudo recente das origens do Homo sapiens geralmente demonstra que existiram outras espécies de Homo, todas as quais estão agora extintas. Enquanto algumas dessas outras espécies poderiam ter sido ancestrais do H. sapiens, muitas foram provavelmente nossos "primos", tendo especificado a partir de nossa linhagem ancestral. Ainda não há nenhum consenso a respeito de quais desses grupos deveriam ser considerados como espécies em separado e sobre quais deveriam ser subespécies de outras espécies. Em alguns casos, isso é devido à escassez de fósseis, em outros, devido a diferenças mínimas usadas para distinguir espécies no gênero Homo. Houve também fluxo gênico entre as diferentes espécies do gênero Homo, o que possibilitou a conservação de alguns genes de espécies extintas no genoma de humanos modernos[113].
H. habilis[editar | editar código-fonte]
Homo habilis é uma espécie de hominídeo que viveu no princípio do Pleistoceno inferior (há 2,2 milhões a 780 mil anos). Os primeiros fósseis de H. habilis foram descobertos em 1964 por Louis Leakey e seus colegas, no desfiladeiro de Olduvai, Tanzânia, que faz parte do Grande Vale do Rift, na África oriental.[114]
Após a descrição da espécie em 1964, H. habilis foi altamente contestado e muitos pesquisadores denominavam essa espécie como um sinônimo do Australopithecus africanus, o único outro hominínio primitivo conhecido na época. Entretanto, o H. habilis recebeu mais reconhecimento com as novas descobertas nos anos posteriores. Na década de 1980, foi proposto que H. habilis foi um ancestral humano, evoluindo diretamente para o Homo erectus, que levou diretamente aos humanos modernos. Este ponto de vista está agora em debate.
As proporções corporais de H. habilis são conhecidas apenas a partir de dois esqueletos altamente fragmentados e se baseiam amplamente na suposição de uma anatomia semelhante aos australopitecos anteriores. Por causa disso, também foi proposto que H. habilis fosse movido para o gênero Australopithecus como Australopithecus habilis.
A espécie Homo habilis apresenta expansão craniana de cerca de 650 cm³ de cérebro, um encurtamento de neurocrânio e um achatamento da parte superior. Além disso, a face apresenta estrutura achatada em relação aos hominínios anteriores. A dentição é parecida com a do H. sapiens com caninos pequenos e dentes achatados.
Poucos fósseis de H. habilis foram encontrados, o que dificulta descrever detalhes precisos da espécie. Porém estes fósseis foram encontrados associados a ferramentas de pedra lascada e por isso essa espécie foi chamada de Homo habilis - homem fabril (ou homem com capacidade de fabricar). Entretanto, hoje sabe-se que essas ferramentas já eram utilizadas anteriormente por outras espécies de hominínios. Além disso, o H. habilis apesar de bípede, apresentava uma bipedia facultativa.[114]H. erectus[editar | editar código-fonte]

Homo erectus é uma espécie extinta de hominínio que evoluiu na África e viveu há entre 1,8 milhões de anos[115] e 100-200 mil anos[116] (Pleistoceno inferior e médio).[117][118] seus espécimes estão entre os primeiros membros reconhecíveis do gênero Homo. H. erectus foi o primeiro ancestral humano a se espalhar pela Eurásia, com uma distribuição continental que se estende da Península Ibérica até a ilha de Java, na Indonésia. É provável que as populações africanas de H. erectus sejam os ancestrais de várias espécies humanas, como H. heidelbergensis e H. antecessor, sendo o primeiro geralmente considerado o ancestral dos neandertais e denisovanos, e às vezes também dos humanos modernos.[119][120][121] Populações asiáticas de H. erectus podem ser ancestrais de H. floresiensis[122] e possivelmente de H. luzonensis.[123] Como uma cronoespécie, a época do desaparecimento do H. erectus é motivo de controvérsia. Existem também várias subespécies propostas com vários níveis de reconhecimento. A última população conhecida de H. erectus é H. e. soloensis de Java, por volta de 117.000–108.000 anos atrás.[124]
H. erectus tinha uma maneira de andar e proporções corporais semelhantes às humanas, e foi a primeira espécie humana a exibir uma face plana, nariz proeminente e, possivelmente, cobertura escassa de pêlos no corpo. Embora o tamanho do cérebro certamente exceda o das espécies ancestrais, a capacidade variava amplamente, dependendo da população. Em populações mais velhas, o desenvolvimento do cérebro parecia ser interrompido cedo na infância, sugerindo que a prole era amplamente autossuficiente ao nascer, limitando assim o desenvolvimento cognitivo ao longo da vida. No entanto, as instalações geralmente mostram o consumo de animais de médio a grande porte, como bovinos ou elefantes, e sugerem o desenvolvimento de comportamento predatório e caça coordenada. H. erectus está associado com a indústria acheuliana de ferramentas de pedra, e postula-se que foi o ancestral humano mais antigo capaz de usar fogo, caçar e coletar em grupos coordenados, cuidar de membros do grupo feridos ou doentes e, possivelmente, navegação e arte (embora sejam exemplos da arte são controversos e, de resto, rudimentares e poucos e distantes entre si).
Homens e mulheres de H. erectus podem ter aproximadamente o mesmo tamanho entre si (isto é, exibiram dimorfismo sexual reduzido) como os humanos modernos, o que poderia indicar monogamia em linha com as tendências gerais exibidas em primatas. O tamanho, no entanto, variou amplamente de 146-185 cm de altura e 40-68 kg de peso. Não está claro se H. erectus era anatomicamente capaz de falar, embora seja postulado que eles se comunicaram usando alguma protolinguagem.H.ergaster[editar | editar código-fonte]

Homo ergaster ou Homo erectus ergaster[125] é uma espécie de hominídeo descrita a partir de restos fossilizados encontrados em Swartkrans, na África do Sul, com uma idade estimada entre 1,8 e um milhão de anos. á grande disputa na comunidade científica sobre se o Homo ergaster é uma espécie em si, ou se a mesma deveria ser introduzida como uma subespécie do Homo erectus, com os defensores do H. Ergaster demarcando que as diferenças entre os fósseis encontrados na África e os da Ásia possuem diferenças significativas o suficiente para indicar uma espécie em si.
Similarmente ao seus antepassados na Garganta de Olduvai, o Homo ergaster possuia grande aptidão na criação e uso de ferramentas, sendo capaz de meticulosamente transformar lascas de rocha em “machados de mão” com bordas afiadas que podiam ser usadas para uma variedade de tarefas. Tal capacidade, assim como seu possível domínio sobre o fogo, ajuda a explicar sinais de fósseis que indicam uma expansão do Homo ergaster para a Eurásia, começando a exploração de novas terras e a chegada desta espécie de hominíneo a climas nunca antes vistosH. heidelbergensis[editar | editar código-fonte]
H. floresiensis[editar | editar código-fonte]

Homo floresiensis (homem de flores), apelidado de "Hobbit", é uma espécie extinta de hominínio do gênero Homo.[126][127]
Os primeiros ossos foram descobertos em 2003 durante uma escavação arqueológica em uma caverna em Liang Bua, Ilha de Flores, na Indonésia.[128] O primeiro fóssil encontrado em Flores foi o esqueleto quase completo de uma mulher, com o crânio por inteiro, referenciado como "LB1". Esse registro tem como característica a baixa estatura, chegando a somente 1 metro de altura, e a baixa capacidade craniana[127] e foi datado para o final do Pleistoceno.[128] Após a primeira escavação, fósseis parciais de outros indivíduos também foram encontrados, confirmando a existência de uma população de pequenos hominínios, e não somente um indivíduo anômalo.[126]
A partir de análises iniciais de carvão da área do primeiro registro, foi concluído que a espécie viveu até 30 mil anos após a chegada de humanos modernos. Porém, escavações mais recentes concluíram que a extinção da espécie ocorreu cerca de 50 mil anos atrás, presumivelmente em razão de dificuldades impostas pelos novos hominíneos na região.[129] Apesar dessa possível ligação, ainda não existem evidências que comprovem interações entre Homo floresiensis e Homo sapiens.[130]
Desde sua descoberta, o status taxonômico do homem de flores causou diversas controvérsias. Muito se debateu se o fóssil encontrado pertencia a uma nova espécie, ou foi causado por um evento patológico. Estudos filogenéticos concluíram que a linhagem do Homo floresiensis compõe efetivamente uma nova espécie, com possível ligação com Homo habilis ou Homo erectus.[131]H. neanderthalensis[editar | editar código-fonte]
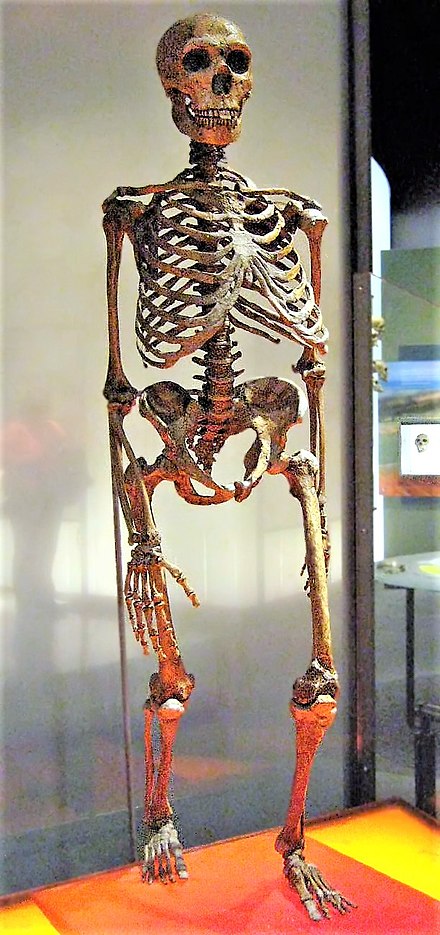
O homem de Neandertal (Homo neanderthalensis) é uma espécie irmã de Homo sapiens, com a qual o homem moderno conviveu.[132][133] Surgiu durante o Pleistoceno Médio na Europa e no Médio Oriente há cerca de 400 mil anos e extinguiu-se há 28 mil anos, na Península Ibérica.[134] As razões para a extinção dos neandertais ainda são debatidas, com diversas possíveis causas levantadas para este fenômeno. As teorias mais aceitas apontam para fatores demográficos como pequeno tamanho populacional e endocruzamento[135], cruzamento e assimilação por humanos modernos[136], mudanças climáticas[137][138][139], doenças e a combinação desses.[140]
O fóssil mais antigo possivelmente pertencente à espécie data de há 430 mil anos.[141] Evidências genéticas indicam que o último ancestral comum (LCA do inglês "Last common ancestor") entre H. neanderthalensis e H. sapiens viveu entre entre 550 e 765 mil anos atrás na África, e seria um representante de Homo heidelbergensis.[142][143][144] Ambas as espécies conviveram após à migração de H. sapiens para a Eurásia, com o primeiro encontro das duas espécies tendo ocorrido a cerca de 80 mil anos no oriente médio, onde hoje se encontra Israel.[145][146]
H. neandertalensis possui grandes porções de seu DNA similares às sequências dos humanos modernos. Além disso, o sequenciamento de DNA nuclear neandertal realizado desde 2006 e publicado a partir de 2010 mostrou um antigo “fluxo gênico” entre os homens neandertais e os homens modernos da Eurásia. Os humanos não africanos atuais têm entre 1,8 e 2,6% dos genes neandertais, adquiridos por hibridação há cerca de 50 mil anos, logo após sua saída da África, e mais de 30% do genoma neandertal sobrevive em toda a população atual em diferentes locais do nosso genoma.[147] Acredita-se também que uma pequena parte desses genes adquiridos por hibridização que estão amplamente presentes em humanos tenham sido selecionados na nossa espécie por trazerem vantagens adaptativas.[148]
Apesar da similaridade genética entre as duas espécies, e da ocorrência de hibridização entre elas, H. neanderthalensis e H. sapiens são espécies distintas que apresentavam diferenças morfológicas significativas.[149] Quando comparados com humanos modernos, os neandertais apresentavam corpos mais robustos com mais reservas de gordura[150] e maiores narizes para o aquecimento do ar[151], ambas características associadas a adaptação ao clima frio[151](a fixação dessas características, no entanto, poderia ter sido resultado de deriva genética[150]). A capacidade craniana de homens e mulheres neandertais era em média de 1600 cm3 e 1300 cm3 respectivamente[152][153], o que é consideravelmente maior do que a média de 1400 e 1200 cm3 de humanos modernos.[154] Além disso, o crânio de neandertais era mais alongado e o cérebro possuía lobos parietais[155] [156] e um cerebelo menor[157], mas com regiões temporais, occipitais e orbitofrontais maiores.[158][159] Progressos relativos da arqueologia pré-histórica e da paleoantropologia, posteriores à década de 1960, descrevem-no como um ser de considerável cultura, mas eventualmente subestimada por alguns autores.[160]
A cultura material e tecnológica dos neandertais era sofisticada em vários aspectos. Os neandertais produziam artefatos de pedra-lascada pertencentes a cultura musteriense, por meio da técnica de levallois.[161] Além de ferramentas, também usavam o fogo[162], eram caçadores de topo[163] e já cuidariam dos doentes.[164][165][166] Há inclusive, quem neles reconheça capacidades estéticas e espirituais semelhantes às do homem atual, como as sugeridas por pingentes[167], pinturas[168][169][170], gravuras[171] e possíveis sepulturas.[172]Homo sapiens[editar | editar código-fonte]

Humano (taxonomicamente Homo sapiens,[173][174] termo que deriva do latim "homem sábio",[175] também conhecido como pessoa, gente ou homem) é a única espécie do gênero Homo ainda viva[173][174] e o primata mais abundante e difundido da Terra, caracterizado pelo bipedalismo e por cérebros grandes, o que permitiu o desenvolvimento de ferramentas, culturas e linguagens avançadas. Os humanos tendem a viver em estruturas sociais complexas compostas por muitos grupos cooperantes e concorrentes, desde famílias e redes de parentesco até Estados políticos. As interações sociais entre os humanos estabeleceram uma ampla variedade de valores, normas e rituais, que fortalecem a sociedade humana. A curiosidade e o desejo humano de compreender e influenciar o meio ambiente e de explicar e manipular fenômenos motivaram o desenvolvimento da ciência, filosofia, mitologia, religião e outros campos de estudo da humanidade.
O H. sapiens surgiu há cerca de 300 mil anos na África, quando evoluiu do Homo heidelbergensis e migrou para fora do continente africano, substituindo gradualmente as populações locais de humanos arcaicos. Durante a maior parte da história, todos os humanos foram caçadores-coletores nômades. A Revolução Neolítica, que começou no sudoeste da Ásia há cerca de 13 mil anos, trouxe o surgimento da agricultura e da ocupação humana permanente. À medida que as populações se tornaram maiores e mais densas, formas de governança se desenvolveram dentro e entre as comunidades e várias civilizações surgiram e declinaram. Os humanos continuaram a se expandir, com uma população global de mais de 8 bilhões em novembro de 2022.[176]
Os genes e o ambiente influenciam a variação biológica humana em características visíveis, fisiologia, suscetibilidade a doenças, habilidades mentais, tamanho do corpo e longevidade. Embora variem em muitas características, dois humanos são, em média, mais de 99% semelhantes. Geralmente, os homens têm maior força corporal, enquanto as mulheres apresentam maior percentual de gordura corporal, entram na menopausa e tornam-se inférteis por volta dos 50 anos e, em média, também têm uma expectativa de vida mais longa em quase todas as populações do mundo. A natureza dos papéis de gênero masculino e feminino tem variado historicamente e os desafios às normas de gênero predominantes têm se repetido em muitas sociedades. Os humanos são onívoros, capazes de consumir uma grande variedade de materiais vegetais e animais e usam o fogo e outras formas de calor para preparar e cozinhar alimentos desde a época do H. erectus. Eles podem sobreviver por até oito semanas sem comida e três ou quatro dias sem água. Geralmente são diurnos, dormindo em média sete a nove horas por dia. O parto é perigoso, com alto risco de complicações e morte. Frequentemente, a mãe e o pai cuidam dos filhos, que são indefesos ao nascer.
Os humanos têm um córtex pré-frontal grande e altamente desenvolvido, a região do cérebro associada à cognição superior. Eles são inteligentes, capazes de memória episódica, expressões faciais flexíveis, autoconsciência, mentalização, introspecção, pensamento privado, imaginação, volição e formação de pontos de vista sobre sua própria existência. Isso tem permitido grandes avanços tecnológicos e o desenvolvimento de ferramentas complexas, possíveis por meio da razão e da transmissão de conhecimento às gerações futuras. Linguagem, arte e comércio são características definidoras dos humanos. As rotas comerciais de longa distância podem ter levado a explosões culturais e distribuição de recursos que deram aos humanos uma vantagem sobre outras espécies semelhantes. A África Oriental, nomeadamente o Chifre da África, é considerada pelos antropólogos como o local de nascimento dos humanos de acordo com as evidências arqueológicas e fósseis existentes.[177][178][179][177]Evolução recente de Homo sapiens[editar | editar código-fonte]
Genética e cultura[editar | editar código-fonte]
Como exemplificado nos tópicos anteriores, muitas pressões seletivas diferentes afetaram a evolução humana. Um desses fatores que vem sendo estudado é o papel da cultura em Homo sapiens.[180] Assim como temos uma herança genética, a herança cultural é bastante marcante em nossa espécie e também pode ser alvo da seleção natural.[181] A Teoria da coevolução gene-cultura é um ramo da genética populacional que estuda justamente como esses dois tipos de transmissão de heranças podem afetar um ao outro e portanto moldar a evolução humana[182]
É possível estudar a coevolução entre genes e cultura por meio de análises genômicas, partindo da premissa de que a cultura tem interferência na evolução humana.[180] Para tal, alguns estudos podem focar em genes candidatos que sofreram seleção positiva nos últimos 100,000 anos.[181] normalmente associados a fenótipos que podem ser atrelados a prática culturais, mas também é possível realizar uma análise a nível genômico, por meio de observações do polimorfismo de nucleotídeo único em diferentes populações, possibilitando inferências sobre as diferentes seleções envolvidas na evolução de fenótipos.[180][183][184] Segundo alguns desses estudos, a transmissão cultural pode estar diretamente ligada a uma seleção sexual, resultando em pressões seletivas locais de características físicas claramente visíveis, como é o caso dos pigmentos da pele e cabelos. Há evidências da teoria também dentro da antropologia e arqueologia.[180] Eles trazem exemplos de como práticas culturais humanas podem mudar o meio e, portanto, as pressões seletivas que sofremos, levando à mudança da frequência de alelos de uma população[185]

Um exemplo é o caso das populações falantes das línguas Cuás, no oeste da África. Dentre uma prática agrícola comum está a abertura de clareiras nas florestas para o cultivo de inhame, o que acarreta em um acúmulo de água da chuva e propicia um ambiente adequado para a reprodução de mosquitos vetores da malária. Essa situação tornou-se uma pressão seletiva para a seleção da hemoglobina S nessas populações, pois indivíduos heterozigotos para os alelos da hemoglobina não contraem a malária. Além disso, foi verificado que em populações próximas, que apresentam práticas agrícolas diferentes dos falantes das línguas Cuás, não foi observado o aumento na frequência alélica da hemoglobina S, reforçando assim a coevolução entre gene-cultura.[180][186]
A investigação do impacto da cultura na nossa evolução se baseia em várias técnicas do estudo da evolução humana.[180] Com o sequenciamento genético dos registros paleontológicos de hominíneos, é possível inferir a história evolutiva dos diferentes grupos associados à evolução humana [187][188]. A partir deles, torna-se possível observar sinais de cruzamentos inter-específicos, tal como ocorreu entre neandertais e hominíneos de Denisova;[188] e também serem considerados complementares aos inúmeros estudos realizados com fósseis[189][190], enriquecendo as inferências e dando suporte às hipóteses atualmente em vigor[188]
Um dos métodos mais promissores para os estudos ligados à história e evolução das populações humanas é o sequenciamento do DNA mitocondrial.[188][191][192][193][194][195][196] O principal motivo para tal está intrinsecamente ligado às propriedades desse material genético, já que apresenta diversas cópias, é transmitido maternalmente, não sofre recombinação genética, é uma molécula pequena, tem uma organização gênica constante e alta taxa de mutações[197][192][195]
A partir das análises mitocondriais, tornou-se possível a reconstrução da origem e história migratória de Homo sapiens ancestrais, utilizando-se de inferências do relógio molecular,[197][193][196] além de possibilitar uma análise epidemiológica acerca de doenças em populações modernas,[193] bem como elucidar a evolução e diversidade genética de povos pouco estudados, como os aborígenes australianos[194][195]
Nesses casos, a busca por sinais de seleção em genomas está amparada por métodos matemáticos e estatísticos, baseados em fatores como desequilíbrio de ligação, análise de haplótipos e variantes raras presentes em grandes frequências. Apesar disso, a interpretação desses dados deve ser cuidadosa e questionada, pois os modelos não são infalíveis e é possível haver algum viés amostral que venha a interferir nas análises.[180][198]
Dentre os diversos genes que já foram estudados, com o foco na evolução recente de Homo sapiens atrelada à cultura, é possível destacar aqueles relacionados com a imunidade, digestão de produtos lácteos, resistência ou quebra de compostos secundários de plantas, metabolismo energético, fenótipos visíveis externamente, desenvolvimento esquelético, fibras musculares da mandíbula e sistema nervoso.[180]
Persistência da lactase[editar | editar código-fonte]

Em conjunto com o surgimento das glândulas mamárias, mamíferos desenvolveram a capacidade de hidrólise da lactose, carboidrato encontrado no leite, para que a absorção de carboidratos menores (galactose e glicose) pelo intestino delgado fosse possível, uma vez que a lactose não é digerida por nosso trato digestivo. A hidrólise realizada pela enzima lactase é essencial para a fase de aleitamento, em que a prole alimenta-se estritamente de leite. Durante o desenvolvimento, essa enzima sofre decréscimo em sua expressão, chegando a taxas muito baixas ao atingir a vida adulta. No entanto, diferentemente de outros mamíferos, humanos apresentam a persistência dessa enzima durante a fase adulta.[199]

Aproximadamente um terço dos seres humanos apresenta, o que denominamos, persistência da lactase (lactase persistence, LP). A frequência da LP varia entre as populações, podendo apresentar taxas de 5% até quase 100%. As maiores frequências são encontradas entre pessoas do nordeste europeu, populações do oeste e leste da África e do Oriente Médio.[199]
A persistência da lactase é um exemplo de evolução convergente, em que diferentes variantes, presentes em populações distintas, convergem para o mesmo fenótipo, nesse caso, a capacidade de produzir lactase na fase adulta. A LP é uma característica autossômica dominante e, sabe-se de ao menos cinco mutações distintas que descrevem esse fenótipo. Evidências sugerem que a seleção positiva da LP ocorreu concomitantemente em diferentes continentes, após a domesticação de animais para ordenha a aproximadamente 10.500 anos atrás, com os primeiros registros no Oriente Médio, espalhando-se para Europa (9 mil anos) e África (7 mil anos).[199]
A correlação entre o consumo de leite e a frequência do fenótipo LP corroboram para a hipótese histórico-cultural. Questiona-se, no entanto, se a capacidade de hidrólise da lactose na vida adulta é anterior ou posterior ao hábito de ingerir leite mesmo depois do desmame. Projeções sugerem que para o surgimento de uma nova mutação em alelos associados à persistência da lactase, seriam necessários 7 mil anos, enquanto a domesticação de animais para a ordenha data pouco mais de 10 mil anos, ou seja, muito tempo para que o leite fosse consumido sem a capacidade de quebrar a lactose. Portanto, as mutações associadas com a LP já deveriam estar presentes em baixas frequências nas populações antes da domesticação para ordenha.[199]
Para a validação da hipótese histórico-cultural questiona-se também qual seria o ganho em aptidão para indivíduos com a persistência da lactase. Uma possível explicação é de que a ingestão de leite seria benéfica do ponto de vista nutricional e energético, fonte de proteína, gordura, minerais e vitaminas. Mesmo pessoas sem a capacidade de hidrolisar lactose poderiam ingerir leite, se em quantidades moderadas e intervaladas durante o dia, a depender da eficiência de fermentação realizada pela microbiota presente no cólon intestinal. Ou mesmo, consumir produtos derivados que possuem menor quantidade de lactose devido sua fermentação, beneficiando-se da glicose presente nesses produtos. No entanto, carboidratos não eram escassos nas populações humanas para que pudesse ser considerada como pressão seletiva, e mesmo em situações de escassez a produção de leite pelo pastoreio não seria viável.[199]
A hipótese da assimilação de cálcio propõe que o leite seria uma fonte importante deste mineral, assim como vitamina D e lactose que auxiliariam na absorção do primeiro. A hipótese faz um recorte para populações que habitam altas latitudes, como é o caso dos europeus, que teriam uma dieta pobre em vitamina D e que também sofrem com a baixa incidência de radiação solar, essencial para a produção dessa vitamina em nossos corpos. No entanto, estudos apontam que pessoas sem o fenótipo LP se beneficiaram mais consumindo produtos derivados do leite para a absorção de cálcio, do que indivíduos de fenótipo LP, divergindo da hipótese de que a lactose auxilia na assimilação desse mineral.[199]
Assim como a hipótese da assimilação de cálcio, a hipótese de que o leite seria uma fonte importante de água e eletrólitos para regiões com escassez de água não contaminada faz um recorte geográfico, compreendendo a África e o Oriente Médio, mas que não explica a ocorrência da persistência da lactase ao redor do globo.[199]
Estudos feitos com uma amostra maior de populações, no entanto, descartam as duas últimas hipóteses, concluindo que a correlação entre consumo de leite e presença de fenótipo LP concorda apenas com a hipótese histórico-cultural, ou seja, com a cultura do pastoreio nas sociedades humanas. Na África a frequência da persistência da lactase é consistente com essa hipótese. No entanto, no cenário europeu o fenótipo LP relacionado a domesticação de animais em conjunto com a facilidade de assimilação do cálcio devido às altas latitudes, ainda apresenta conflitos.[199]
Analisando as frequências sem a distinção das localidades, estudos encontraram uma correlação mundial de alta significância entre o pastoreio e a LP. No entanto, separando as frequências para cada continente africano, europeu e asiático, respectivamente, notou-se que a África é a responsável por forçar essa correlação, uma vez que a correlação não é significante para a Europa, para a Ásia ou Eurásia.[199]
Adaptação ao mergulho em Homo sapiens[editar | editar código-fonte]
Como mencionado no tópico Genética e Cultura existe uma forte correlação entre cultura e genética passível de estudos de coevolução.[180] Diversos estudos buscam elucidar a relação entre esses dois assuntos e a maioria faz uso de exemplos atuais, como a persistência da lactase e presença de hemoglobina S em populações do oeste da África. Outro exemplo notável são as populações nômades do mar do sudeste asiático que possuem hábitos de mergulho há milhares de anos e apresentam alterações fenotípicas decorrentes desse hábito[200]

As sociedades nômades dos mares do sudeste da China são populações que, desde o período da pré-história até os dias atuais, vivem migrando entre os arquipélagos do que hoje conhecemos como Taiwan, Malásia, Filipinas e Indonésia.[201][202] Esses grupos de nômades do mar tinham hábitos caçadores-coletores, sem indícios de agricultura,[202] e contribuíram para o estabelecimento de rotas de trocas de mercadorias por todo o oceano do sudeste asiático, passando pelo oeste da África e chegando à Oceania. As trocas não se resumiam a mercadorias, mas também envolviam ideias, culturas e pessoas, através de toda a região Indo-Pacífica.[201] Esses povos são originários de aproximadamente 3000 anos antes da era comum, de uma migração de Taiwan conhecida como expansão Austronesiana (tradução livre de Austronesian expansion)[203]

Dentre esses povos se encontram os Sama bajau, que compreendem mais de 1 milhão de pessoas entre Filipinas, Indonésia e Malásia. Os Bajaus são um grupo étnico dentro dos Austronésios, que habitam a região e são exímios mergulhadores caçadores, vivendo na costa de diversas ilhas do sudeste asiático em grupos fragmentados.[203] Esses mergulhadores apresentam capacidade de mergulho superior comparativamente a outros povos não mergulhadores da mesma região, pois são capazes de prender a respiração por longos períodos de tempo. Além disso, apresentam baço aumentado, de modo que armazenam mais sangue do que outras pessoas, o que significa mais oxigênio durante mergulhos de profundidade.
Diversos genes candidatos podem estar envolvidos com essas adaptações ao mergulho, como por exemplo o gene PDE10A, que está relacionado com a contração da musculatura lisa, incluindo da região do baço, ou então o gene BDKRB2, o qual evidências sugerem forte seleção nos Bajaus, e afeta o reflexo de mergulho dos humanos. Podemos mencionar também o gene FAM178B, que codifica uma proteína que forma um complexo estável com a anidrase carbônica, auxiliando na manutenção do pH do sangue.[200]
De modo geral os Bajaus passaram por adaptações únicas com relação a suas respostas de mergulho, exemplificando uma adaptação humana notável na história evolutiva recente. Eles e outras populações mergulhadoras representam uma nova oportunidade de estudo de adaptações humanas à hipóxia, da mesma forma que sociedades andinas, que apresentam quantidades superiores de hemoglobina para suportar o ar rarefeito.[200] Além disso, da mesma forma que populações com adaptações de dieta decorrentes do pastoralismo, essas adaptações com base genética surgiram como consequência de novas práticas culturais, ilustrando que a cultura e a genética tem co-evoluido por milhares de anos[200]
Evolução da arte[editar | editar código-fonte]
A presença da arte é vista de alguma forma em todas as culturas humanas, sendo muitas vezes definida como um traço universal humano [204]. Assim, quando olhamos o perfil arqueológico, evidências de arte são uma das evidências chaves para indicar a atividade humana moderna[205], e fornecer informações sobre desenvolvimento cognitivo e a evolução cultural[206].
O surgimento da arte visual tem sido hipotetizado por ter acontecido durante a era paleolítica, por volta de 40.000 anos atrás, no qual aconteceu uma explosão criativa. No entanto, evidências recentes sugerem que ocorria processamento de pigmento e produção de ornamentos na Era da Pedra Média Africana, datando pelo menos a 120000 anos atrás[207]. Independente do momento exato do seu surgimento, a partir do início da atividade artística, presume-se que os humanos adquiriram a habilidade de pensar em símbolos e figurativamente, permitindo a habilidade de se expressar artisticamente. Tal avanço cognitivo é utilizado para estimar o surgimento da linguagem nos humanos[208].
A Evolução da arte hominínea pode ser utilizada para inferir o desenvolvimento da evolução cognitiva dos hominídeos ao longo do tempo, uma vez que a presença de arte envolve a presença de cultura e pode possuir importante papel na comunicação, mediando informação e expressão.[209]
Ver também[editar | editar código-fonte]
- ↑ a b Fuentes, Agustín (fevereiro de 2015). «Integrative Anthropology and the Human Niche: Toward a Contemporary Approach to Human Evolution: Integrative Anthropology and the Human Niche». American Anthropologist (em inglês) (2): 302–315. doi:10.1111/aman.12248. Consultado em 15 de julho de 2022
- ↑ Heng HH (2009). «The genome-centric concept: resynthesis of evolutionary theory». Bioessays. 31 (5): 512–25. PMID 19334004. doi:10.1002/bies.200800182
- ↑ Stringer, C.B. (1994). «Evolution of early humans». In: Steve Jones, Robert Martin & David Pilbeam (eds.). The Cambridge Encyclopedia of Human Evolution. Cambridge: Cambridge University Press. p. 242. ISBN 0-521-32370-3 Also ISBN 0-521-46786-1 (paperback)
- ↑ McHenry, H.M (2009). «Human Evolution». In: Michael Ruse & Joseph Travis. Evolution: The First Four Billion Years. Cambridge, Massachusetts: The Belknap Press of Harvard University Press. p. 265. ISBN 978-0-674-03175-3
- ↑ «Out of Africa Revisited». Science. 13 de maio de 2005. Consultado em 9 de março de 2017
- ↑ Chris Stringer (12 de junho de 2003). «Human evolution: Out of Ethiopia». Nature. Consultado em 9 de março de 2017
- ↑ «Origins of Modern Humans: Multiregional or Out of Africa?». actionbioscience. Maio de 2001. Consultado em 9 de março de 2017
- ↑ Afinal foi na Europa? Fóssil com 8.7 milhões de anos coloca em causa a origem dos humanos, ZAP, 25 de Acosto de 2023
- ↑ Brues & Snow 1965, pp. 1–39
- ↑ a b c d e f Da Silva, Sergio Francisco Serafim Monteiro (1 de fevereiro de 2020). «ASSIM CAMINHOU A HUMANIDADE». CLIO Arqueológica (3). 171 páginas. ISSN 2448-2331. doi:10.20891/clio.v34n3p171-193. Consultado em 17 de julho de 2021
- ↑ a b Schmitt, Daniel (1 de maio de 2003). «Insights into the evolution of human bipedalism from experimental studies of humans and other primates». Journal of Experimental Biology (9): 1437–1448. ISSN 1477-9145. doi:10.1242/jeb.00279. Consultado em 17 de julho de 2021
- ↑ a b DART, RAYMOND A. (fevereiro de 1925). «Australopithecus africanus The Man-Ape of South Africa». Nature (2884): 195–199. ISSN 0028-0836. doi:10.1038/115195a0. Consultado em 17 de julho de 2021
- ↑ a b Maslin, Mark A.; Shultz, Susanne; Trauth, Martin H. (5 de março de 2015). «A synthesis of the theories and concepts of early human evolution». Philosophical Transactions of the Royal Society B: Biological Sciences (1663). 20140064 páginas. ISSN 0962-8436. doi:10.1098/rstb.2014.0064. Consultado em 17 de julho de 2021
- ↑ a b c d e f g h Ko, Kwang Hyun (dezembro de 2015). «Origins of Bipedalism». Brazilian Archives of Biology and Technology (6): 929–934. ISSN 1516-8913. doi:10.1590/S1516-89132015060399. Consultado em 17 de julho de 2021
- ↑ White, T. D.; Asfaw, B.; Beyene, Y.; Haile-Selassie, Y.; Lovejoy, C. O.; Suwa, G.; Woldegabriel, G. (2009). «Ardipithecus ramidus and the Paleobiology of Early Hominids». Science. 326 (5949): 75–86. Bibcode:2009Sci...326...75W. PMID 19810190. doi:10.1126/science.1175802
- ↑ Wayman, Erin (16 de março de 2012). «Data Deluge Drives Demand». Science. ISSN 1095-9203. doi:10.1126/science.caredit.a1200030. Consultado em 17 de julho de 2021
- ↑ a b c d e f Niemitz, Carsten (março de 2010). «The evolution of the upright posture and gait—a review and a new synthesis». Naturwissenschaften (em inglês) (3): 241–263. ISSN 0028-1042. PMC 2819487
. PMID 20127307. doi:10.1007/s00114-009-0637-3. Consultado em 17 de julho de 2021
- ↑ Darwin, Charles (1874). The descent of man : and selection in relation to sex / By Charles Darwin. New York :: Merrill and Baker
- ↑ a b Kramer, P.A.; Eck, G.G. (maio de 2000). «Locomotor energetics and leg length in hominid bipedality». Journal of Human Evolution (em inglês) (5): 651–666. doi:10.1006/jhev.1999.0375. Consultado em 17 de julho de 2021
- ↑ Sockol, Michael D.; Raichlen, David A.; Pontzer, Herman (24 de julho de 2007). «Chimpanzee locomotor energetics and the origin of human bipedalism». Proc. Natl. Acad. Sci. U.S.A. 104 (30): 12265–12269. Bibcode:2007PNAS..10412265S. ISSN 0027-8424. PMC 1941460
- «Study identifies energy efficiency as reason for evolution of upright walking». UANews. Tucson, AZ: The University of Arizona Office of University Communications. 16 de julho de 2007. Consultado em 23 de abril de 2015
- ↑ a b Kramer, Patricia; Sylvester, Adam (2009). «Bipedal Form and Locomotor Function: Understanding the Effects of Size and Shape on Velocity and Energetics». PaleoAnthropology. ISSN 1545-0031. doi:10.4207/pa.2009.art32. Consultado em 17 de julho de 2021
- ↑ a b Hardy, Alister (1960). «Was man more aquatic in the past» (PDF). New scientist (em inglês). 7 (642): 5. Consultado em 15 de junho de 2021. Cópia arquivada (PDF) em 26 de março de 2009
- ↑ Langdon, John H. (outubro de 1997). «Umbrella hypotheses and parsimony in human evolution: a critique of the Aquatic Ape Hypothesis». Journal of Human Evolution (4): 479–494. ISSN 0047-2484. doi:10.1006/jhev.1997.0146. Consultado em 17 de julho de 2021
- ↑ Rose, M. D. (março de 1976). «Bipedal behavior of olive baboons (Papio anubis) and its relevance to an understanding of the evolution of human bipedalism». American Journal of Physical Anthropology (2): 247–261. ISSN 0002-9483. doi:10.1002/ajpa.1330440207. Consultado em 17 de julho de 2021
- ↑ a b c Hunt, Kevin D. (março de 1994). «The evolution of human bipedality: ecology and functional morphology». Journal of Human Evolution (3): 183–202. ISSN 0047-2484. doi:10.1006/jhev.1994.1011. Consultado em 17 de julho de 2021
- ↑ Jolly, Clifford J. (março de 1970). «The Seed-Eaters: A New Model of Hominid Differentiation Based on a Baboon Analogy». Man (1). 5 páginas. doi:10.2307/2798801. Consultado em 17 de julho de 2021
- ↑ Wrangham, R.W. (maio de 1980). «Bipedal locomotion as a feeding adaptation in gelada baboons, and its implications for hominid evolution». Journal of Human Evolution (4): 329–331. ISSN 0047-2484. doi:10.1016/0047-2484(80)90059-7. Consultado em 17 de julho de 2021
- ↑ WARD, E. J.; UNDERWOOD, C. R. (julho de 1967). «The Effect of Posture on the Solar Radiation Area of Man». Ergonomics (4): 399–409. ISSN 0014-0139. doi:10.1080/00140136708930887. Consultado em 17 de julho de 2021
- ↑ a b Sussman, R. W.; Hart, Donna. «The Behavioral Ecology of our Earliest Hominid Ancestors». New York, NY: Springer New York: 259–279. ISBN 978-0-387-73895-6. Consultado em 17 de julho de 2021
- ↑ a b Kwang Hyun, Ko (2015). «Origins of Bipedalism». Brazilian Archives of Biology and Technology. 58 (6): 929–934. Bibcode:2015arXiv150802739K. arXiv:1508.02739
- ↑ DeSalle & Tattersall 2008, p. 113
- ↑ a b Curry 2008, pp. 106–109
- ↑ David-Barrett, T.; Dunbar, R.I.M. (2016). «Bipedality and Hair-loss Revisited: The Impact of Altitude and Activity Scheduling». Journal of Human Evolution. 94: 72–82. PMC 4874949
- ↑ Harcourt-Smith, W. E. H.; Aiello, L. C. (maio de 2004). «Fossils, feet and the evolution of human bipedal locomotion». Journal of Anatomy (em inglês) (5): 403–416. ISSN 0021-8782. PMC 1571304
- ↑ Srivastava, R.P. (2009). Morphology of the Primates and Human Evolution. New Delhi: PHI Learning Private Limited. ISBN 978-81-203-3656-8. OCLC 423293609
- ↑ Strickberger 2000, pp. 475–476
- ↑ Trevathan 2011, p. 20
- ↑ Zuk, Marlene (2014), "Paleofantasy: What Evolution Really Tells Us About Sex, Diet, and How We Live" (W.W. Norton & Company)
- ↑ Hrdy, Sarah Blaffer (2011), "Mothers and Others: The Evolutionary Origins of Mutual Understanding" (Harvard Uni Press)
- ↑ Wayman, Erin (19 de agosto de 2013). «Killer whales, grandmas and what men want: Evolutionary biologists consider menopause». Science News. ISSN 0036-8423. Consultado em 24 de abril de 2015
- ↑ Zollikofer, Christoph P. E.; Ponce de León, Marcia S.; Lieberman, Daniel E.; Guy, Franck; Pilbeam, David; Likius, Andossa; Mackaye, Hassane T.; Vignaud, Patrick; Brunet, Michel (abril de 2005). «Virtual cranial reconstruction of Sahelanthropus tchadensis». Nature (em inglês) (7034): 755–759. ISSN 1476-4687. doi:10.1038/nature03397. Consultado em 19 de julho de 2021
- ↑ a b Schoenemann, P. Thomas (Outubro de 2006). «Evolution of the Size and Functional Areas of the Human Brain». Annual Review of Anthropology. 35: 379–406. ISSN 0084-6570. doi:10.1146/annurev.anthro.35.081705.123210
- ↑ Brown, Graham; Fairfax, Stephanie; Sarao, Nidhi. «Tree of Life Web Project: Human Evolution». www.tolweb.org
- ↑ Park, Min S.; Nguyen, Andrew D.; Aryan, Henry E.; et al. (Março de 2007). «Evolution of the human brain: changing brain size and the fossil record». Neurosurgery. 60 (3): 555–562. ISSN 0148-396X. PMID 17327801. doi:10.1227/01.NEU.0000249284.54137.32
- ↑ Bruner, Emiliano (Dezembro de 2007). «Cranial shape and size variation in human evolution: structural and functional perspectives». Child's Nervous System. 23 (12): 1357–1365. CiteSeerX 10.1.1.391.288
- ↑ Potts, Richard (Outubro de 2012). «Evolution and Environmental Change in Early Human Prehistory». Annual Review of Anthropology. 41: 151–167. ISSN 0084-6570. doi:10.1146/annurev-anthro-092611-145754
- ↑ Leonard, William R.; Snodgrass, J. Josh; Robertson, Marcia L. (Agosto de 2007). «Effects of brain evolution on human nutrition and metabolism». Annual Review of Nutrition. 27: 311–327. ISSN 0199-9885. PMID 17439362. doi:10.1146/annurev.nutr.27.061406.093659
- ↑ McBroom, Patricia (14 de junho de 1999). «Meat-eating was essential for human evolution, says UC Berkeley anthropologist specializing in diet» (Nota de imprensa). Berkeley: University of California Press. Consultado em 25 de abril de 2015
- ↑ Mann, Neil (Setembro de 2007). «Meat in the human diet: An anthropological perspective». Nutrition & Dietetics. 64 (Supplement s4): S102–S107. ISSN 1747-0080. doi:10.1111/j.1747-0080.2007.00194.x. Consultado em 31 de janeiro de 2012
- ↑ Zimmer, Carl (13 de agosto de 2015). «For Evolving Brains, a 'Paleo' Diet Full of Carbs». The New York Times. Consultado em 14 de agosto de 2015
- ↑ Organ, Chris; Nunn, Charles L.; Machanda, Zarin; Wrangham, Richard W. (30 de agosto de 2011). «Phylogenetic rate shifts in feeding time during the evolution of Homo». Proc. Natl. Acad. Sci. U.S.A. 108 (35): 14555–14559. Bibcode:2011PNAS..10814555O. ISSN 0027-8424. PMC 3167533
- ↑ David-Barrett, T.; Dunbar, R.I.M. (2013). «Processing Power Limits Social Group Size: Computational Evidence for the Cognitive Costs of Sociality». Proceedings of the Royal Society B. 280 (1765): 20131151. PMC 3712454
- ↑ Bown & Rose 1987
- ↑ Barton, Robert A.; Venditti, Chris (20 de Outubro de 2014). «Rapid Evolution of the Cerebellum in Humans and Other Great Apes». Current Biology. 24 (20): 2440–2444. ISSN 0960-9822. PMID 25283776. doi:10.1016/j.cub.2014.08.056
- ↑ Starowicz-Filip, Anna; Milczarek, Olga; Kwiatkowski, Stanisław; et al. (2013). «Cerebellar cognitive affective syndrome CCAS – a case report». Archives of Psychiatry and Psychotherapy. 15 (3): 57–64. OCLC 220954272. doi:10.12740/APP/18666
- ↑ Feng Yu; Qing-jun Jiang; Xi-yan Sun; et al. (22 de agosto de 2014). «A new case of complete primary cerebellar agenesis: clinical and imaging findings in a living patient». Brain. 138 (Pt 6): e353. ISSN 1460-2156. PMC 4614135
- ↑ Weaver, Anne H. (8 de março de 2005). «Reciprocal evolution of the cerebellum and neocortex in fossil humans». Proc. Natl. Acad. Sci. U.S.A. 102 (10): 3576–3580. Bibcode:2005PNAS..102.3576W. ISSN 0027-8424. PMC 553338
- ↑ Hauser, Marc D.; Chomsky, Noam; Fitch, W. Tecumseh (22 de novembro de 2002). «The Faculty of Language: What Is It, Who Has It, and How Did It Evolve?». Science (em inglês) (5598): 1569–1579. ISSN 0036-8075. doi:10.1126/science.298.5598.1569. Consultado em 12 de julho de 2022
- ↑ Klein, Stefan (2014). Survival of the Nicest. [S.l.]: The Experiment. ISBN 978-1615190904
- ↑ Sallet, J.; Mars, R.B.; Noonan, M.P.; Andersson, J.L.; O'Reilly, J.X.; Jbabdi, S.; Croxson, P.L.; Jenkinson, M.; Miller, K.L. (4 de novembro de 2011). «Social Network Size Affects Neural Circuits in Macaques». Science. 334 (6056): 697–700. Bibcode:2011Sci...334..697S. ISSN 0036-8075. PMID 22053054. doi:10.1126/science.1210027
- ↑ Dunbar, R.I.M. (1992). «Neocortex size as a constraint on group size in primates». Journal of Human Evolution. 22 (6): 469–493. doi:10.1016/0047-2484(92)90081-j
- ↑ Dávid-Barrett, T.; Dunbar, R.I.M. (22 de agosto de 2013). «Processing power limits social group size: computational evidence for the cognitive costs of sociality». Proceedings of the Royal Society of London B: Biological Sciences. 280 (1765). 20131151 páginas. ISSN 0962-8452. PMC 3712454
- ↑ Shultz, Susanne; Dunbar, Robin (14 de dezembro de 2010). «Encephalization is not a universal macroevolutionary phenomenon in mammals but is associated with sociality». Proceedings of the National Academy of Sciences. 107 (50): 21582–21586. Bibcode:2010PNAS..10721582S. ISSN 0027-8424. PMC 3003036
- ↑ Richard, Potts (1998). «Environmental Hypotheses of Hominin Evolution». American Journal of Physical Anthropology. 107: 93–136. doi:10.1002/(SICI)1096-8644(1998)107:27 (inativo 6 de dezembro de 2019)
- ↑ Kaplan, Hillard; Hill, Kim; Lancaster, Jane; Hurtado, Magdelena (16 de agosto de 2000). «A Theory of Human Life History Evolution: Diet, Intelligence, and Longevity» (PDF). Evolutionary Anthropology. 9 (4): 156–185. doi:10.1002/1520-6505(2000)9:4<156::AID-EVAN5>3.0.CO;2-7
- ↑ Samal, Ashok; Subramani, Vanitha; Marx, David (dezembro de 2007). «Analysis of sexual dimorphism in human face». Journal of Visual Communication and Image Representation (6): 453–463. ISSN 1047-3203. doi:10.1016/j.jvcir.2007.04.010. Consultado em 17 de julho de 2021
- ↑ a b Reno, Philip L.; Lovejoy, C. Owen (28 de abril de 2015). «From Lucy to Kadanuumuu: balanced analyses of Australopithecus afarensisassemblages confirm only moderate skeletal dimorphism». PeerJ: e925. ISSN 2167-8359. doi:10.7717/peerj.925. Consultado em 17 de julho de 2021
- ↑ Lovejoy, C. O. (1 de outubro de 2009). «Reexamining Human Origins in Light of Ardipithecus ramidus». Science (5949): 74–74, 74e1-74e8. ISSN 0036-8075. doi:10.1126/science.1175834. Consultado em 17 de julho de 2021
- ↑ a b c d «Sex, Reproduction, and Scenarios of Human Evolution». The MIT Press. 2018. ISBN 978-0-262-34418-0. Consultado em 17 de julho de 2021
- ↑ a b c d e f Wells, Jonathan C.K. (setembro de 2007). «Sexual dimorphism of body composition». Best Practice & Research Clinical Endocrinology & Metabolism (3): 415–430. ISSN 1521-690X. doi:10.1016/j.beem.2007.04.007. Consultado em 17 de julho de 2021
- ↑ Wiskott, L.; Fellous, J.-M.; Kuiger, N.; von der Malsburg, C. (julho de 1997). «Face recognition by elastic bunch graph matching». IEEE Transactions on Pattern Analysis and Machine Intelligence (7): 775–779. ISSN 0162-8828. doi:10.1109/34.598235. Consultado em 17 de julho de 2021
- ↑ Maynard, L. M.; Wisemandle, W.; Roche, A. F.; Chumlea, Wm. C.; Guo, S. S.; Siervogel, R. M. (1 de fevereiro de 2001). «Childhood Body Composition in Relation to Body Mass Index». PEDIATRICS (2): 344–350. ISSN 0031-4005. doi:10.1542/peds.107.2.344. Consultado em 17 de julho de 2021
- ↑ Zamboni, Mauro; Zoico, Elena; Scartezzini, Tiziana; Mazzali, Gloria; Tosoni, Paolo; Zivelonghi, Alessandra; Gallagher, Dympna; De Pergola, Giovanni; Di Francesco, Vincenzo (agosto de 2003). «Body composition changes in stable-weight elderly subjects: The effect of sex». Aging Clinical and Experimental Research (4): 321–327. ISSN 1594-0667. doi:10.1007/bf03324517. Consultado em 17 de julho de 2021
- ↑ Darwin, Charles (1871). The descent of man, and selection in relation to sex. By Charles Darwin ... New York,: D. Appleton and company,
- ↑ Puts, David Andrew; Gaulin, Steven J.C.; Verdolini, Katherine (julho de 2006). «Dominance and the evolution of sexual dimorphism in human voice pitch». Evolution and Human Behavior (4): 283–296. ISSN 1090-5138. doi:10.1016/j.evolhumbehav.2005.11.003. Consultado em 17 de julho de 2021
- ↑ Feinberg, David R.; Jones, Benedict C.; Armstrong, Marie M. (dezembro de 2018). «Sensory Exploitation, Sexual Dimorphism, and Human Voice Pitch». Trends in Ecology & Evolution (12): 901–903. ISSN 0169-5347. doi:10.1016/j.tree.2018.09.007. Consultado em 17 de julho de 2021
- ↑ Rosenbaum, M. (1 de junho de 1999). «Role of Gonadal Steroids in the Sexual Dimorphisms in Body Composition and Circulating Concentrations of Leptin». Journal of Clinical Endocrinology & Metabolism (6): 1784–1789. ISSN 0021-972X. doi:10.1210/jc.84.6.1784. Consultado em 17 de julho de 2021
- ↑ Young, Richard W (janeiro de 2003). «Evolution of the human hand: the role of throwing and clubbing». Journal of Anatomy. 202 (1): 165–174. ISSN 0021-8782. PMC 1571064
- ↑ a b Miriam Ittyerah (2013). Hand Preference and Hand Ability: Evidence from studies in Haptic Cognition. [S.l.]: John Benjamins Publishing. pp. 37–38. ISBN 978-9027271648
- ↑ Frank R. Wilson. «The Hand How Its Use Shapes the Brain, Language, and Human Culture». The New York Times. Pantheon Books. Consultado em 2 de julho de 2017
- ↑ Pontzer, Herman (2016). «Metabolic acceleration and the evolution of human brain size and life history». Nature. 533 (7603): 390–392. Bibcode:2016Natur.533..390P. PMC 4942851
- ↑ a b Seyfarth, Robert M.; Cheney, Dorothy L. (fevereiro de 2017). «Precursors to language: Social cognition and pragmatic inference in primates». Psychonomic Bulletin & Review (em inglês) (1): 79–84. ISSN 1069-9384. doi:10.3758/s13423-016-1059-9. Consultado em 26 de junho de 2024
- ↑ Aboitiz, Francisco; Garcı́a V., Ricardo (dezembro de 1997). «The evolutionary origin of the language areas in the human brain. A neuroanatomical perspective». Brain Research Reviews (3): 381–396. ISSN 0165-0173. doi:10.1016/s0165-0173(97)00053-2. Consultado em 27 de junho de 2024
- ↑ Schenker, N. M.; Hopkins, W. D.; Spocter, M. A.; Garrison, A. R.; Stimpson, C. D.; Erwin, J. M.; Hof, P. R.; Sherwood, C. C. (20 de julho de 2009). «Broca's Area Homologue in Chimpanzees (Pan troglodytes): Probabilistic Mapping, Asymmetry, and Comparison to Humans». Cerebral Cortex (3): 730–742. ISSN 1047-3211. PMC PMC2820707
|pmc=(ajuda). PMID 19620620. doi:10.1093/cercor/bhp138. Consultado em 27 de junho de 2024 - ↑ Aboitiz, Francisco; Garcı́a V., Ricardo (dezembro de 1997). «The evolutionary origin of the language areas in the human brain. A neuroanatomical perspective». Brain Research Reviews (3): 381–396. ISSN 0165-0173. doi:10.1016/s0165-0173(97)00053-2. Consultado em 27 de junho de 2024
- ↑ a b c d e f g Zhang, Elizabeth Qing; Shi, Edward Ruoyang; Barceló-Coblijn, Lluís (27 de abril de 2023). «Categorical perception and language evolution: a comparative and neurological perspective». Frontiers in Psychology (em English). ISSN 1664-1078. PMC PMC10172646
|pmc=(ajuda). PMID 37179894 Verifique|pmid=(ajuda). doi:10.3389/fpsyg.2023.1110730. Consultado em 26 de junho de 2024 - ↑ Hockett, Charles F. (setembro de 1960). «The Origin of Speech». Scientific American (3): 88–96. ISSN 0036-8733. doi:10.1038/scientificamerican0960-88. Consultado em 26 de junho de 2024
- ↑ a b c d e Engesser, Sabrina; Townsend, Simon W. (julho de 2019). «Combinatoriality in the vocal systems of nonhuman animals». WIREs Cognitive Science (em inglês) (4): e1493. ISSN 1939-5078. doi:10.1002/wcs.1493. Consultado em 26 de junho de 2024
- ↑ Bowling, Daniel L.; Fitch, W. Tecumseh (outubro de 2015). «Do Animal Communication Systems Have Phonemes?». Trends in Cognitive Sciences (10): 555–557. ISSN 1364-6613. doi:10.1016/j.tics.2015.08.011. Consultado em 26 de junho de 2024
- ↑ Rohrmeier, Martin; Zuidema, Willem; Wiggins, Geraint A.; Scharff, Constance (19 de março de 2015). «Principles of structure building in music, language and animal song». Philosophical Transactions of the Royal Society B: Biological Sciences (1664). 20140097 páginas. ISSN 0962-8436. doi:10.1098/rstb.2014.0097. Consultado em 26 de junho de 2024
- ↑ Hauser, Marc D.; Chomsky, Noam; Fitch, W. Tecumseh (7 de janeiro de 2010). «The faculty of language: what is it, who has it, and how did it evolve?». Cambridge University Press: 14–42. ISBN 978-0-521-51645-7. Consultado em 26 de junho de 2024
- ↑ Costa, Raquel (Janeiro, 2014). «Uma ponte entre a aprendizagem social e cultura em primatas não humanos.»
- ↑ Davidson, Iain (2016). «Stone Tools: Evidence of Something in Between Culture and Cumulative Culture?». Dordrecht: Springer Netherlands: 99–120. ISBN 978-94-017-7424-6. Consultado em 14 de julho de 2022
- ↑ Maxwell 1984, p. 296
- ↑ Rui Zhang; Yin-Qiu Wang; Bing Su (julho de 2008). «Molecular Evolution of a Primate-Specific microRNA Family» (PDF). Molecular Biology and Evolution. 25 (7): 1493–1502. ISSN 0737-4038. PMID 18417486. doi:10.1093/molbev/msn094. Consultado em 27 de abril de 2015
- ↑ Willoughby, Pamela R. (2005). «Palaeoanthropology and the Evolutionary Place of Humans in Nature». International Journal of Comparative Psychology. 18 (1): 60–91. ISSN 0889-3667. Consultado em 27 de abril de 2015
- ↑ Martin 2001, pp. 12032–12038
- ↑ Tavaré, Simon; Marshall, Charles R.; Will, Oliver; et al. (18 de abril de 2002). «Using the fossil record to estimate the age of the last common ancestor of extant primates». Nature. 416 (6882): 726–729. Bibcode:2002Natur.416..726T. ISSN 0028-0836. PMID 11961552. doi:10.1038/416726a
- ↑ Rose, Kenneth D. (1994). «The earliest primates». Evolutionary Anthropology: Issues, News, and Reviews. 3 (5): 159–173. ISSN 1060-1538. doi:10.1002/evan.1360030505
- ↑ Fleagle, John; Gilbert, Chris (2011). Rowe, Noel; Myers, Marc, eds. «Primate Evolution». All The World's Primates. Charlestown, RI: Primate Conservation, Inc. Consultado em 27 de abril de 2015
- ↑ Roach, John (3 de março de 2008). «Oldest Primate Fossil in North America Discovered». National Geographic News. Washington, DC: National Geographic Society. Consultado em 27 de abril de 2015
- ↑ McMains, Vanessa (5 de dezembro de 2011). «Found in Wyoming: New fossils of oldest American primate». The Gazette. Baltimore, MD: Johns Hopkins University. Consultado em 27 de abril de 2015
- ↑ Caldwell, Sara B. (19 de maio de 2009). «Missing link found, early primate fossil 47 million years old». Digital Journal. Toronto, Canada: digitaljournal.com. Consultado em 27 de abril de 2015
- ↑ Watts, Alex (20 de maio de 2009). «Scientists Unveil Missing Link In Evolution». Sky News Online. London: BSkyB. Consultado em 27 de abril de 2015. Cópia arquivada em 28 de julho de 2011
- ↑ Wilford, J.N. (5 de junho de 2013). «Palm-size fossil resets primates' clock, scientists say». The New York Times. Consultado em 5 de junho de 2013
- ↑ Kordos, László; Begun, David R. (janeiro de 2001). «Primates from Rudabánya: allocation of specimens to individuals, sex and age categories». Journal of Human Evolution. 40 (1): 17–39. ISSN 0047-2484. PMID 11139358. doi:10.1006/jhev.2000.0437
- ↑ Cameron 2004, p. 76
- ↑ Wallace 2004, p. 240
- ↑ Zalmout, Iyad S.; Sanders, William J.; MacLatchy, Laura M.; et al. (15 de julho de 2010). «New Oligocene primate from Saudi Arabia and the divergence of apes and Old World monkeys». Nature. 466 (7304): 360–364. Bibcode:2010Natur.466..360Z. ISSN 0028-0836. PMID 20631798. doi:10.1038/nature09094
- ↑ Srivastava 2009, p. 87
- ↑ Clark, G.; Henneberg, M. (junho de 2015). «The life history of Ardipithecus ramidus: a heterochronic model of sexual and social maturation». Anthropological Review. 78 (2): 109–132. doi:10.1515/anre-2015-0009. Consultado em 22 de julho de 2015
- ↑ Clark, G.; Henneberg, M. (June 2015). "The life history of Ardipithecus ramidus: a heterochronic model of sexual and social maturation". Anthropological Review. 78 (2): 109–132. doi:10.1515/anre-2015-0009. Retrieved 2015-07-22.
- ↑ Kuhlwilm, Martin; Gronau, Ilan; Hubisz, Melissa J.; de Filippo, Cesare; Prado-Martinez, Javier; Kircher, Martin; Fu, Qiaomei; Burbano, Hernán A.; Lalueza-Fox, Carles (fevereiro de 2016). «Ancient gene flow from early modern humans into Eastern Neanderthals». Nature (em inglês) (7591): 429–433. ISSN 0028-0836. PMC PMC4933530
|pmc=(ajuda). PMID 26886800. doi:10.1038/nature16544. Consultado em 5 de julho de 2023 - ↑ a b Faria, Felipe (2014). Memória: o homem habilidoso, Revista Ciência Hoje (SBPC), n.311, janeiro 2014. [S.l.: s.n.] ISSN 0101-8515
- ↑ Redação - avph.com.br. «Homo erectus». avph.com.br. Consultado em 23 de junho de 2011. Cópia arquivada em 23 de fevereiro de 2015
- ↑ Herries, Andy I. R.; Martin, Jesse M.; Leece, A. B.; Adams, Justin W.; Boschian, Giovanni; Joannes-Boyau, Renaud; Edwards, Tara R.; Mallett, Tom; Massey, Jason (3 de abril de 2020). «Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa». Science (em inglês). 368 (6486). ISSN 0036-8075. PMID 32241925. doi:10.1126/science.aaw7293
- ↑ Redação - Infopédia. «Homo erectus». Infopédia. Consultado em 23 de junho de 2011. Cópia arquivada em 23 de fevereiro de 2015
- ↑ Herries, Andy I. R.; Martin, Jesse M.; Leece, A. B.; Adams, Justin W.; Boschian, Giovanni; Joannes-Boyau, Renaud; Edwards, Tara R.; Mallett, Tom; Massey, Jason (3 de abril de 2020). «Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa». Science (em inglês) (6486). ISSN 0036-8075. PMID 32241925. doi:10.1126/science.aaw7293. Consultado em 20 de julho de 2021
- ↑ Klein, R. (1999). The Human Career: Human Biological and Cultural Origins. Chicago: University of Chicago Press
- ↑ Wood, Bernard (28 de junho de 2011). «Did early Homo migrate "out of" or "in to" Africa?». Proceedings of the National Academy of Sciences (26): 10375–10376. Consultado em 20 de julho de 2021
- ↑ Ko, Kwang Hyun (16 de julho de 2016). «Hominin interbreeding and the evolution of human variation». Journal of Biological Research-Thessaloniki (1). 17 páginas. ISSN 2241-5793. PMC 4947341
- ↑ van den Bergh, Gerrit D.; Kaifu, Yousuke; Kurniawan, Iwan; Kono, Reiko T.; Brumm, Adam; Setiyabudi, Erick; Aziz, Fachroel; Morwood, Michael J. (1 de junho de 2016). «Homo floresiensis-like fossils from the early Middle Pleistocene of Flores». Nature: 245–248. ISSN 0028-0836. doi:10.1038/nature17999. Consultado em 20 de julho de 2021
- ↑ Détroit, Florent; Mijares, Armand Salvador; Corny, Julien; Daver, Guillaume; Zanolli, Clément; Dizon, Eusebio; Robles, Emil; Grün, Rainer; Piper, Philip J. (abril de 2019). «A new species of Homo from the Late Pleistocene of the Philippines». Nature (em inglês) (7751): 181–186. ISSN 1476-4687. doi:10.1038/s41586-019-1067-9. Consultado em 20 de julho de 2021
- ↑ Rizal, Y.; Westaway, K.; Zaim, Y.; Bergh, G. van den; Bettis, E.; Morwood, M.; Huffman, O. F.; Grün, R.; Joannes-Boyau, R. (2019). «Last appearance of Homo erectus at Ngandong, Java, 117,000–108,000 years ago». Nature. doi:10.1038/s41586-019-1863-2. Consultado em 20 de julho de 2021
- ↑ Noel T. Boaz, Russell L. Ciochon, Dragon Bone Hill: An Ice-Age Saga of Homo erectus, Oxford University Press (2004).
- ↑ a b Morwood, M. J.; Brown, P.; Jatmiko; Sutikna, T.; Wahyu Saptomo, E.; Westaway, K. E.; Awe Due, Rokus; Roberts, R. G.; Maeda, T. (outubro de 2005). «Further evidence for small-bodied hominins from the Late Pleistocene of Flores, Indonesia». Nature (7061): 1012–1017. ISSN 0028-0836. doi:10.1038/nature04022. Consultado em 15 de julho de 2021
- ↑ a b Neves, Walter; Diniz-Filho, José Alexandre Felizola (julho de 2018). «O Hobbit da Ilha de Flores: implicações para a evolução humana». Ciência e Cultura (3): 56–59. ISSN 2317-6660. doi:10.21800/2317-66602018000300015. Consultado em 15 de julho de 2021
- ↑ a b Brown, P.; Sutikna, T.; Morwood, M. J.; Soejono, R. P.; Jatmiko; Wayhu Saptomo, E.; Awe Due, Rokus (outubro de 2004). «A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia». Nature (7012): 1055–1061. ISSN 0028-0836. doi:10.1038/nature02999. Consultado em 15 de julho de 2021
- ↑ Callaway, Ewen (30 de março de 2016). «Did humans drive 'hobbit' species to extinction?». Nature. ISSN 0028-0836. doi:10.1038/nature.2016.19651. Consultado em 15 de julho de 2021
- ↑ Sutikna, Thomas; Tocheri, Matthew W.; Morwood, Michael J.; Saptomo, E. Wahyu; Jatmiko; Awe, Rokus Due; Wasisto, Sri; Westaway, Kira E.; Aubert, Maxime (30 de março de 2016). «Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia». Nature (7599): 366–369. ISSN 0028-0836. doi:10.1038/nature17179. Consultado em 15 de julho de 2021
- ↑ Argue, Debbie; Groves, Colin P.; Lee, Michael S.Y.; Jungers, William L. (junho de 2017). «The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters». Journal of Human Evolution: 107–133. ISSN 0047-2484. doi:10.1016/j.jhevol.2017.02.006. Consultado em 15 de julho de 2021
- ↑ «A surpreendente forma como o homem de Neanderthal pegou herpes - BBC Brasil». Consultado em 2 de agosto de 2016
- ↑ «Dentro de cada humano mora um neandertal | VEJA.com». Consultado em 2 de agosto de 2016
- ↑ Finlayson, Clive; Giles Pacheco, Francisco; Rodríguez-Vidal, Joaquín; Fa, Darren A.; María Gutierrez López, José; Santiago Pérez, Antonio; Finlayson, Geraldine; Allue, Ethel; Baena Preysler, Javier (outubro de 2006). «Late survival of Neanderthals at the southernmost extreme of Europe». Nature (em inglês) (7113): 850–853. ISSN 1476-4687. doi:10.1038/nature05195. Consultado em 5 de julho de 2023
- ↑ Vaesen, Krist; Dusseldorp, Gerrit L.; Brandt, Mark J. (1 de março de 2021). «An emerging consensus in palaeoanthropology: demography was the main factor responsible for the disappearance of Neanderthals». Scientific Reports (em inglês) (1). ISSN 2045-2322. PMC PMC7921565
|pmc=(ajuda). PMID 33649483. doi:10.1038/s41598-021-84410-7. Consultado em 5 de julho de 2023 - ↑ Smith, Fred H. (setembro de 2011). «Assimilation Revisited: Africans, Neandertals, and the Origins of Modern Eurasians». General Anthropology (em inglês) (2): 1–7. doi:10.1111/j.1939-3466.2011.00009.x. Consultado em 5 de julho de 2023
- ↑ Bradtmöller, Marcel; Pastoors, Andreas; Weninger, Bernhard; Weniger, Gerd-Christian (janeiro de 2012). «The repeated replacement model – Rapid climate change and population dynamics in Late Pleistocene Europe». Quaternary International (em inglês): 38–49. doi:10.1016/j.quaint.2010.10.015. Consultado em 5 de julho de 2023
- ↑ Wolf, D.; Kolb, T.; Alcaraz-Castaño, M.; Heinrich, S.; Baumgart, P.; Calvo, R.; Sánchez, J.; Ryborz, K.; Schäfer, I. (4 de maio de 2018). «Climate deteriorations and Neanderthal demise in interior Iberia». Scientific Reports (em inglês) (1). ISSN 2045-2322. doi:10.1038/s41598-018-25343-6. Consultado em 5 de julho de 2023
- ↑ Black, Benjamin A.; Neely, Ryan R.; Manga, Michael (maio de 2015). «Campanian Ignimbrite volcanism, climate, and the final decline of the Neanderthals». Geology (em inglês) (5): 411–414. ISSN 1943-2682. doi:10.1130/G36514.1. Consultado em 5 de julho de 2023
- ↑ Timmermann, Axel (junho de 2020). «Quantifying the potential causes of Neanderthal extinction: Abrupt climate change versus competition and interbreeding». Quaternary Science Reviews (em inglês). 106331 páginas. doi:10.1016/j.quascirev.2020.106331. Consultado em 5 de julho de 2023
- ↑ Arsuaga, J. L.; Martínez, I.; Arnold, L. J.; Aranburu, A.; Gracia-Téllez, A.; Sharp, W. D.; Quam, R. M.; Falguères, C.; Pantoja-Pérez, A. (20 de junho de 2014). «Neandertal roots: Cranial and chronological evidence from Sima de los Huesos». Science (em inglês) (6190): 1358–1363. ISSN 0036-8075. doi:10.1126/science.1253958. Consultado em 5 de julho de 2023
- ↑ Meyer, Matthias; Arsuaga, Juan-Luis; de Filippo, Cesare; Nagel, Sarah; Aximu-Petri, Ayinuer; Nickel, Birgit; Martínez, Ignacio; Gracia, Ana; de Castro, José María Bermúdez (março de 2016). «Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins». Nature (em inglês) (7595): 504–507. ISSN 1476-4687. doi:10.1038/nature17405. Consultado em 5 de julho de 2023
- ↑ Mounier, A.; Caparros, M. (1 de outubro de 2015). «Le statut phylogénétique d'Homo heidelbergensis – étude cladistique des homininés du Pléistocène moyen». BMSAP (em francês) (3): 110–134. ISSN 1777-5469. doi:10.1007/s13219-015-0127-4. Consultado em 5 de julho de 2023
- ↑ Stringer, Chris (maio de 2012). «The status of Homo heidelbergensis (Schoetensack 1908)». Evolutionary Anthropology: Issues, News, and Reviews (em inglês) (3): 101–107. doi:10.1002/evan.21311. Consultado em 5 de julho de 2023
- ↑ Higham, Tom; Douka, Katerina; Wood, Rachel; Ramsey, Christopher Bronk; Brock, Fiona; Basell, Laura; Camps, Marta; Arrizabalaga, Alvaro; Baena, Javier (21 de agosto de 2014). «The timing and spatiotemporal patterning of Neanderthal disappearance». Nature (em inglês) (7514): 306–309. ISSN 0028-0836. doi:10.1038/nature13621. Consultado em 5 de julho de 2023
- ↑ Gibbons, A. (7 de maio de 2010). «Close Encounters of the Prehistoric Kind». Science (em inglês) (5979): 680–684. ISSN 0036-8075. doi:10.1126/science.328.5979.680. Consultado em 5 de julho de 2023
- ↑ Spector, Dina. «Here's What Happened When Neanderthals And Ancient Humans Hooked Up 80,000 Years Ago»
- ↑ Singer, Emily. «How Neanderthal DNA Helps Humanity» (em inglês)
- ↑ «Homo neanderthalensis». The Smithsonian Institution's Human Origins Program (em inglês). Consultado em 5 de julho de 2023
- ↑ a b Weaver, Timothy D. (22 de setembro de 2009). «The meaning of Neandertal skeletal morphology». Proceedings of the National Academy of Sciences (em inglês) (38): 16028–16033. ISSN 0027-8424. doi:10.1073/pnas.0903864106. Consultado em 5 de julho de 2023
- ↑ a b de Azevedo, S.; González, M. F.; Cintas, C.; Ramallo, V.; Quinto-Sánchez, M.; Márquez, F.; Hünemeier, T.; Paschetta, C.; Ruderman, A. (21 de novembro de 2017). «Nasal airflow simulations suggest convergent adaptation in Neanderthals and modern humans». Proceedings of the National Academy of Sciences (em inglês) (47): 12442–12447. ISSN 0027-8424. PMC PMC5703271
|pmc=(ajuda). PMID 29087302. doi:10.1073/pnas.1703790114. Consultado em 5 de julho de 2023 - ↑ Foley, Robert, ed. (1984). Hominid evolution and community ecology: prehistoric human adaptation in biological perspective. Col: Studies in archaeology. London: Academic Press
- ↑ Amano, Hideki; Kikuchi, Takeo; Morita, Yusuke; Kondo, Osamu; Suzuki, Hiromasa; Ponce de León, Marcia S.; Zollikofer, Christoph P.E.; Bastir, Markus; Stringer, Chris (outubro de 2015). «Virtual reconstruction of the Neanderthal Amud 1 cranium: VIRTUAL RECONSTRUCTION OF AMUD 1 CRANIUM». American Journal of Physical Anthropology (em inglês) (2): 185–197. doi:10.1002/ajpa.22777. Consultado em 5 de julho de 2023
- ↑ Beals, Kenneth L.; Smith, Courtland L.; Dodd, Stephen M.; Angel, J. Lawrence; Armstrong, Este; Blumenberg, Bennett; Girgis, Fakhry G.; Turkel, Spencer; Gibson, Kathleen R. (junho de 1984). «Brain Size, Cranial Morphology, Climate, and Time Machines [and Comments and Reply]». Current Anthropology (em inglês) (3): 301–330. ISSN 0011-3204. doi:10.1086/203138. Consultado em 5 de julho de 2023
- ↑ Bruner, Emiliano (novembro de 2004). «Geometric morphometrics and paleoneurology: brain shape evolution in the genus Homo». Journal of Human Evolution (em inglês) (5): 279–303. doi:10.1016/j.jhevol.2004.03.009. Consultado em 5 de julho de 2023
- ↑ Bruner, Emiliano (junho de 2010). «Morphological Differences in the Parietal Lobes within the Human Genus: A Neurofunctional Perspective». Current Anthropology (em inglês) (S1): S77–S88. ISSN 0011-3204. doi:10.1086/650729. Consultado em 5 de julho de 2023
- ↑ Weaver, Anne H. (8 de março de 2005). «Reciprocal evolution of the cerebellum and neocortex in fossil humans». Proceedings of the National Academy of Sciences (em inglês) (10): 3576–3580. ISSN 0027-8424. doi:10.1073/pnas.0500692102. Consultado em 5 de julho de 2023
- ↑ Bruner, Emiliano; Ogihara, Naomichi; Tanabe, Hiroki C. (28 de dezembro de 2017). Digital Endocasts: From Skulls to Brains (em inglês). [S.l.]: Springer
- ↑ Nishiaki, Yoshihiro; Jöris, Olaf (27 de novembro de 2019). Learning Among Neanderthals and Palaeolithic Modern Humans: Archaeological Evidence (em inglês). [S.l.]: Springer Nature
- ↑ Os neandertais poderão ter sido os primeiros artistas das cavernas – artigo de Ana Gershenfeld (Arte Paleolítica) no jornal Público, Ciência, de 15 de junho de 2012
- ↑ Shaw, Ian, ed. (1999). A dictionary of archaeology. Oxford, UK Malden, Mass: Blackwell Publishers
- ↑ Brittingham, Alex; Hren, Michael T.; Hartman, Gideon; Wilkinson, Keith N.; Mallol, Carolina; Gasparyan, Boris; Adler, Daniel S. (25 de outubro de 2019). «Geochemical Evidence for the Control of Fire by Middle Palaeolithic Hominins». Scientific Reports (em inglês) (1). ISSN 2045-2322. PMC PMC6814844
|pmc=(ajuda). PMID 31653870. doi:10.1038/s41598-019-51433-0. Consultado em 5 de julho de 2023 - ↑ Richards, Michael P.; Pettitt, Paul B.; Trinkaus, Erik; Smith, Fred H.; Paunović, Maja; Karavanić, Ivor (20 de junho de 2000). «Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes». Proceedings of the National Academy of Sciences (em inglês) (13): 7663–7666. ISSN 0027-8424. PMC PMC16602
|pmc=(ajuda). PMID 10852955. doi:10.1073/pnas.120178997. Consultado em 5 de julho de 2023 - ↑ Weyrich, Laura S.; Duchene, Sebastian; Soubrier, Julien; Arriola, Luis; Llamas, Bastien; Breen, James; Morris, Alan G.; Alt, Kurt W.; Caramelli, David (abril de 2017). «Neanderthal behaviour, diet, and disease inferred from ancient DNA in dental calculus». Nature (em inglês) (7650): 357–361. ISSN 1476-4687. doi:10.1038/nature21674. Consultado em 5 de julho de 2023
- ↑ Hardy, Karen; Buckley, Stephen; Huffman, Michael (1 de setembro de 2013). «Neanderthal self-medication in context». Antiquity (em inglês) (337): 873–878. ISSN 0003-598X. doi:10.1017/S0003598X00049528. Consultado em 5 de julho de 2023
- ↑ Spikins, Penny; Needham, Andy; Wright, Barry; Dytham, Calvin; Gatta, Maurizio; Hitchens, Gail (1 de agosto de 2019). «Living to fight another day: The ecological and evolutionary significance of Neanderthal healthcare». Quaternary Science Reviews. Neanderthals: Ecology and Evolution (em inglês): 98–118. ISSN 0277-3791. doi:10.1016/j.quascirev.2018.08.011. Consultado em 5 de julho de 2023
- ↑ Radovčić, Davorka; Sršen, Ankica Oros; Radovčić, Jakov; Frayer, David W. (11 de março de 2015). «Evidence for Neandertal Jewelry: Modified White-Tailed Eagle Claws at Krapina». PLOS ONE (3): e0119802. ISSN 1932-6203. doi:10.1371/journal.pone.0119802. Consultado em 5 de julho de 2023
- ↑ Hoffmann, D. L.; Standish, C. D.; García-Diez, M.; Pettitt, P. B.; Milton, J. A.; Zilhão, J.; Alcolea-González, J. J.; Cantalejo-Duarte, P.; Collado, H. (23 de fevereiro de 2018). «U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art». Science (em inglês) (6378): 912–915. ISSN 0036-8075. doi:10.1126/science.aap7778. Consultado em 5 de julho de 2023
- ↑ Slimak, Ludovic; Fietzke, Jan; Geneste, Jean-Michel; Ontañón, Roberto (21 de setembro de 2018). «Comment on "U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art"». Science (em inglês) (6408). ISSN 0036-8075. doi:10.1126/science.aau1371. Consultado em 5 de julho de 2023
- ↑ White, Randall; Bosinski, Gerhard; Bourrillon, Raphaëlle; Clottes, Jean; Conkey, Margaret W.; Rodriguez, Soledad Corchón; Cortés-Sánchez, Miguel; de la Rasilla Vives, Marco; Delluc, Brigitte (1 de julho de 2020). «Still no archaeological evidence that Neanderthals created Iberian cave art». Journal of Human Evolution (em inglês). 102640 páginas. ISSN 0047-2484. doi:10.1016/j.jhevol.2019.102640. Consultado em 5 de julho de 2023
- ↑ Leder, Dirk; Hermann, Raphael; Hüls, Matthias; Russo, Gabriele; Hoelzmann, Philipp; Nielbock, Ralf; Böhner, Utz; Lehmann, Jens; Meier, Michael (setembro de 2021). «A 51,000-year-old engraved bone reveals Neanderthals' capacity for symbolic behaviour». Nature Ecology & Evolution (em inglês) (9): 1273–1282. ISSN 2397-334X. doi:10.1038/s41559-021-01487-z. Consultado em 5 de julho de 2023
- ↑ Rendu, William; Beauval, Cédric; Crevecoeur, Isabelle; Bayle, Priscilla; Balzeau, Antoine; Bismuth, Thierry; Bourguignon, Laurence; Delfour, Géraldine; Faivre, Jean-Philippe (7 de janeiro de 2014). «Evidence supporting an intentional Neandertal burial at La Chapelle-aux-Saints». Proceedings of the National Academy of Sciences (em inglês) (1): 81–86. ISSN 0027-8424. PMC PMC3890882
|pmc=(ajuda). PMID 24344286. doi:10.1073/pnas.1316780110. Consultado em 5 de julho de 2023 - ↑ a b Goodman et al. 1990, pp. 260–266
- ↑ a b «Hominidae Classification». Animal Diversity Web @ UMich. Consultado em 25 de setembro de 2006. Arquivado do original em 4 de fevereiro de 2004
- ↑ hŏmo, săpĭens, săpĭo. Charlton T. Lewis e Charles Short. A Latin Dictionary em Perseus Project.
- ↑ «World Population». Worldometer (em inglês). Consultado em 17 de novembro de 2022
- ↑ a b Cavalazzi et al. 2019, pp. 553–578
- ↑ Maslin 2017
- ↑ Sahle et al. 2013, p. e78092
- ↑ a b c d e f g h i Laland, Kevin N.; Odling-Smee, John; Myles, Sean (fevereiro de 2010). «How culture shaped the human genome: bringing genetics and the human sciences together». Nature Reviews Genetics (em inglês) (2): 137–148. ISSN 1471-0056. doi:10.1038/nrg2734. Consultado em 19 de julho de 2021
- ↑ a b Voight, Benjamin F.; Kudaravalli, Sridhar; Wen, Xiaoquan; Pritchard, Jonathan K. (7 de março de 2006). «A Map of Recent Positive Selection in the Human Genome». PLOS Biology (em inglês) (3): e72. ISSN 1545-7885. PMC 1382018
- ↑ «Gene-culture coevolutionary theory». Trends in Ecology & Evolution (em inglês) (11): 453–457. 1 de novembro de 1996. ISSN 0169-5347. doi:10.1016/0169-5347(96)10052-5. Consultado em 19 de julho de 2021
- ↑ Lao, O.; de Gruijter, J. M.; van Duijn, K.; Navarro, A.; Kayser, M. (maio de 2007). «Signatures of Positive Selection in Genes Associated with Human Skin Pigmentation as Revealed from Analyses of Single Nucleotide Polymorphisms: Signatures of Positive Selection at Human Pigmentation Genes». Annals of Human Genetics (em inglês) (3): 354–369. doi:10.1111/j.1469-1809.2006.00341.x. Consultado em 19 de julho de 2021
- ↑ Myles, Sean; Somel, Mehmet; Tang, Kun; Kelso, Janet; Stoneking, Mark (19 de dezembro de 2006). «Identifying genes underlying skin pigmentation differences among human populations». Human Genetics (em inglês) (5): 613–621. ISSN 0340-6717. doi:10.1007/s00439-006-0256-4. Consultado em 19 de julho de 2021
- ↑ Ghiselin, Michael T. (fevereiro de 1993). «Coevolution: Genes, Culture, and Human Diversity - William H. Durham, Stanford, CA:Stanford University Press,1991, 629 pp. US$69.50 cloth. ISBN 0-8047-1537-8. Stanford University Press, Stanford University, Stanford, CA 94305, USA.». Politics and the Life Sciences (1): 123–124. ISSN 0730-9384. doi:10.1017/s0730938400011436. Consultado em 19 de julho de 2021
- ↑ Durham, William H. (1991). Coevolution: Genes, Culture, and Human Diversity (em inglês). [S.l.]: Stanford University Press
- ↑ «Something old, something borrowed: admixture and adaptation in human evolution». Current Opinion in Genetics & Development (em inglês): 1–8. 1 de dezembro de 2018. ISSN 0959-437X. doi:10.1016/j.gde.2018.05.009. Consultado em 19 de julho de 2021
- ↑ a b c d Stoneking, Mark (1993). «DNA and recent human evolution». Evolutionary Anthropology: Issues, News, and Reviews (em inglês) (2): 60–73. ISSN 1520-6505. doi:10.1002/evan.1360020208. Consultado em 19 de julho de 2021
- ↑ «Reconstructing recent human evolution». Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences (em inglês) (1280): 217–224. 29 de agosto de 1992. ISSN 0962-8436. doi:10.1098/rstb.1992.0099. Consultado em 19 de julho de 2021
- ↑ Brown, Peter; Aitken, Martin Jim; Stringer, Christopher Brian; Mellars, P. A. (29 de agosto de 1992). «Recent human evolution in East Asia and Australasia». Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences (1280): 235–242. doi:10.1098/rstb.1992.0101. Consultado em 19 de julho de 2021
- ↑ Gray, Michael W. (novembro de 1989). «Origin and Evolution of Mitochondrial DNA». Annual Review of Cell Biology (em inglês) (1): 25–50. ISSN 0743-4634. doi:10.1146/annurev.cb.05.110189.000325. Consultado em 19 de julho de 2021
- ↑ a b Wilson, Allan C.; Cann, Rebecca L.; Carr, Steven M.; George, Matthew; Gyllensten, Ulf B.; Helm-Bychowski, Kathleen M.; Higuchi, Russell G.; Palumbi, Stephen R.; Prager, Ellen M. (dezembro de 1985). «Mitochondrial DNA and two perspectives on evolutionary genetics». Biological Journal of the Linnean Society (em inglês) (4): 375–400. doi:10.1111/j.1095-8312.1985.tb02048.x. Consultado em 19 de julho de 2021
- ↑ a b c Wallace, Douglas C.; Brown, Michael D.; Lott, Marie T. (setembro de 1999). «Mitochondrial DNA variation in human evolution and disease». Gene (em inglês) (1): 211–230. doi:10.1016/S0378-1119(99)00295-4. Consultado em 19 de julho de 2021
- ↑ a b Nagle, Nano; Ballantyne, Kaye N; van Oven, Mannis; Tyler-Smith, Chris; Xue, Yali; Wilcox, Stephen; Wilcox, Leah; Turkalov, Rust; van Oorschot, Roland A H (1 de dezembro de 2016). «Mitochondrial DNA diversity of present-day Aboriginal Australians and implications for human evolution in Oceania». Journal of Human Genetics (3): 343–353. ISSN 1434-5161. doi:10.1038/jhg.2016.147. Consultado em 19 de julho de 2021
- ↑ a b c Nesheva, DV (11 de dezembro de 2014). «Aspects of Ancient Mitochondrial DNA Analysis in Different Populations for Understanding Human Evolution». Balkan Journal of Medical Genetics (1): 5–14. ISSN 1311-0160. PMC 4347471
- ↑ a b Templeton, Alan R. (2007). «Genetics and Recent Human Evolution». Evolution (em inglês) (7): 1507–1519. ISSN 1558-5646. doi:10.1111/j.1558-5646.2007.00164.x. Consultado em 19 de julho de 2021
- ↑ a b Erro de citação: Etiqueta
<ref>inválida; não foi fornecido texto para as refs de nome:13 - ↑ Akey, J. M. (1 de maio de 2009). «Constructing genomic maps of positive selection in humans: Where do we go from here?». Genome Research (5): 711–722. ISSN 1088-9051. doi:10.1101/gr.086652.108. Consultado em 19 de julho de 2021
- ↑ a b c d e f g h i Johnson, Norman (30 de agosto de 2017). «Faculty Opinions recommendation of On the evolution of lactase persistence in humans.». Faculty Opinions – Post-Publication Peer Review of the Biomedical Literature. Consultado em 19 de julho de 2021
- ↑ a b c d «Physiological and Genetic Adaptations to Diving in Sea Nomads». Cell (em inglês) (3): 569–580.e15. 19 de abril de 2018. ISSN 0092-8674. doi:10.1016/j.cell.2018.03.054. Consultado em 19 de julho de 2021
- ↑ a b Kusuma, Pradiptajati; Brucato, Nicolas; Cox, Murray P.; Letellier, Thierry; Manan, Abdul; Nuraini, Chandra; Grangé, Philippe; Sudoyo, Herawati; Ricaut, François-Xavier (agosto de 2017). «The last sea nomads of the Indonesian archipelago: genomic origins and dispersal». European Journal of Human Genetics (em inglês) (8): 1004–1010. ISSN 1476-5438. doi:10.1038/ejhg.2017.88. Consultado em 19 de julho de 2021
- ↑ a b Chen, Jonas Chung-yu (24 de janeiro de 2008). «[ARCHAEOLOGY IN CHINA AND TAIWAN] Sea nomads in prehistory on the southeast coast of China». Bulletin of the Indo-Pacific Prehistory Association (0). ISSN 1835-1794. doi:10.7152/bippa.v22i0.11805. Consultado em 19 de julho de 2021
- ↑ a b Sather, Clifford (setembro de 2006). «Sea Nomads and Rainforest Hunter-Gatherers: Foraging Adaptations in the Indo-Malaysian Archipelago». ANU Press. ISBN 978-0-7315-2132-6. Consultado em 19 de julho de 2021
- ↑ Davidson, I. (2012). Symbolism and becoming a hunter-gatherer. Bull. de la Société Préhistorique Ariège-Pyrénées, 65, 1689.
- ↑ Wadley, L. (2001). What is cultural modernity? A general view and a South African perspective from Rose Cottage Cave. Cambridge Archaeological Journal, 11(2), 201- 221.
- ↑ d'Errico, F., Henshilwood, C., Lawson, G., Vanhaeren, M., Tillier, A.-M., Soressi, M., et al. (2003). Archaeological evidence for the emergence of language, symbolism, and music–an alternative multidisciplinary perspective. Journal of World Prehistory, 17(1), 1-70.
- ↑ McDermott, A. (2021) What was the first “art”? How would we know?. Proceedings of the National Academy of Sciences, 118 (44) e2117561118; DOI: 10.1073/pnas.2117561118
- ↑ Mithen, S.J. (1996). The prehistory of the mind: the cognitive origins of art, religion and science: Thames and Hudson.
- ↑ Nowell, A. (2010). Defining behavioral modernity in the context of Neandertal and anatomically modern human populations. Annual review of anthropology, 39, 437-452.
Bibliografia[editar | editar código-fonte]
- Aiello, Leslie; Dean, Christopher (1990). An Introduction to Human Evolutionary Anatomy. London; San Diego: Elsevier Academic Press. ISBN 978-0-12-045591-1. LCCN 95185095. OCLC 33408268
- Bogin, Barry (1997). «The Evolution of Human Nutrition». In: Romanucci-Ross, Lola; Moerman, Daniel E.; Tancredi, Laurence R. The Anthropology of Medicine: From Culture to Method 3ª ed. Westport, CT: Bergin & Garvey. ISBN 978-0-89789-516-3. LCCN 96053993. OCLC 36165190
- Bown, Thomas M.; Rose, Kenneth D. (1987). Patterns of Dental Evolution in Early Eocene Anaptomorphine Primates (Omomyidae) From the Bighorn Basin, Wyoming. Col: Memoir (Paleontological Society). 23. Tulsa, OK: Paleontological Society. OCLC 16997265. Consultado em 25 de abril de 2015. Arquivado do original (PDF) em 11 de novembro de 2016
- Boyd, Robert; Silk, Joan B. (2003). How Humans Evolved 3ª ed. New York: W.W. Norton & Company. ISBN 978-0-393-97854-4. LCCN 2002075336. OCLC 49959461 (9ª edição, 2021)
- Boyd, Robert; Silk, Joan B. (2021). How Humans Evolved 9ª ed. New York: W.W. Norton & Company. ISBN 978-0-393-53315-6. OCLC 1282924892
- Brues, Alice M.; Snow, Clyde C. (1965). «Physical Anthropology». In: Siegel, Bernard J. Biennial Review of Anthropology 1965. Col: Biennial Review of Anthropology. 4. Stanford, CA: Stanford University Press. ISBN 978-0-8047-1746-5. ISSN 0067-8503. LCCN 59012726. OCLC 01532912
- Bryson, Bill (2004). «The Mysterious Biped». A Short History of Nearly Everything. Toronto: Anchor Canada. ISBN 978-0-385-66004-4. LCCN 2003046006. OCLC 55016591
- Cameron, David W. (2004). Hominid Adaptations and Extinctions. Sydney, NSW: UNSW Press. ISBN 978-0-86840-716-6. LCCN 2004353026. OCLC 57077633
- Cochran, Gregory; Harpending, Henry (2009). The 10,000 Year Explosion: How Civilization Accelerated Human Evolution. New York: Basic Books. ISBN 978-0-465-00221-4. LCCN 2008036672. OCLC 191926088
- Curry, James R. (2008). Children of God: Children of Earth. Bloomington, IN: AuthorHouse. ISBN 978-1-4389-1846-4. OCLC 421466369
- Darwin, Charles (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life (PDF) 1ª ed. [S.l.: s.n.] Consultado em 13 de maio de 2017. Arquivado do original (PDF) em 4 de março de 2016
- Darwin, Charles (1981). The Descent of Man, and Selection in Relation to Sex. Introduction by John Tyler Bonner and Robert M. May republicação ed. Princeton, NJ: Princeton University Press. ISBN 978-0-691-02369-4. LCCN 80008679. OCLC 7197127
- Dawkins, Richard (2004). The Ancestor's Tale: A Pilgrimage to the Dawn of Evolution. Boston, MA: Houghton Mifflin. ISBN 978-0-618-00583-3. LCCN 2004059864. OCLC 56617123
- DeSalle, Rob; Tattersall, Ian (2008). Human Origins: What Bones and Genomes Tell Us About Ourselves. Col: Texas A&M University Anthropology Series. 13 1ª ed. College Station, TX: Texas A&M University Press. ISBN 978-1-58544-567-7. OCLC 144520427
- Diamond, Jared (1999). Guns, Germs, and Steel: The Fates of Human Societies. New York: W.W. Norton & Company. ISBN 978-0-393-31755-8. LCCN 2005284124. OCLC 35792200
- Freeman, Scott; Herron, Jon C. (2007). Evolutionary Analysis 4ª ed. Upper Saddle River, NJ: Pearson Prentice Hall. ISBN 978-0-13-227584-2. LCCN 2006034384. OCLC 73502978
- Johanson, Donald; Edey, Maitland (1981). Lucy, the Beginnings of Humankind. St Albans: Granada. ISBN 978-0-586-08437-3
- Montgomery, William M. (1988). «Germany». In: Glick, Thomas F. The Comparative Reception of Darwinism. Chicago, Illinois: University of Chicago Press. ISBN 978-0-226-29977-8. LCCN 87035814. OCLC 17328115
- Kondo, Shiro, ed. (1985). Primate Morphophysiology, Locomotor Analyses, and Human Bipedalism. American Journal of Physical Anthropology. 70. Tokyo: University of Tokyo Press. pp. 278–279. ISBN 978-4-13-066093-8. LCCN 85173489. OCLC 12352830. doi:10.1002/ajpa.1330700214
- Leakey, Richard E. (1994). The Origin of Humankind. Col: Science Masters Series. New York: Basic Books. ISBN 978-0-465-03135-1. LCCN 94003617. OCLC 30739453
- M'charek, Amade (2005). The Human Genome Diversity Project: An Ethnography of Scientific Practice. Col: Cambridge Studies in Society and the Life Sciences. Cambridge; New York: Cambridge University Press. ISBN 978-0-521-83222-9. LCCN 2004052648. OCLC 55600894
- Martin, Robert D. (2001). «Primates, Evolution of». In: Smelser, Neil J.; Baltes, Paul B. International Encyclopedia of the Social & Behavioral Sciences 1ª ed. Amsterdam; New York: Elsevier. pp. 12032–12038. ISBN 978-0-08-043076-8. LCCN 2001044791. OCLC 47869490. doi:10.1016/B0-08-043076-7/03083-7
- Maxwell, Mary (1984). Human Evolution: A Philosophical Anthropology. New York: Columbia University Press. ISBN 978-0-231-05946-6. LCCN 83024005. OCLC 10163036
- McHenry, Henry M. (2009). «Human Evolution». In: Ruse, Michael; Travis, Joseph. Evolution: The First Four Billion Years. Foreword by Edward O. Wilson. Cambridge, Massachusetts: Belknap Press of Harvard University Press. ISBN 978-0-674-03175-3. LCCN 2008030270. OCLC 225874308
- Ramachandran, Sohini; Hua Tang; Gutenkunst, Ryan N.; Bustamante, Carlos D. (2010). «Genetics and Genomics of Human Population Structure». In: Speicher, Michael R.; Antonarakis, Stylianos E.; Motulsky, Arno G. Vogel and Motulsky's Human Genetics: Problems and Approaches 4ª ed. Heidelberg; London; New York: Springer. ISBN 978-3-540-37653-8. LCCN 2009931325. OCLC 549541244. doi:10.1007/978-3-540-37654-5
- Robinson, J.T. (2008). «Adaptive Radiation in the Australopithecines and the Origin of Man». In: Howell, F. Clark; Bourlière, François. African Ecology and Human Evolution. New Brunswick, NJ: Transaction Publishers. ISBN 978-0-202-36136-9. LCCN 2007024716. OCLC 144770218
- Srivastava, R.P. (2009). Morphology of the Primates and Human Evolution. New Delhi: PHI Learning Private Limited. ISBN 978-81-203-3656-8. OCLC 423293609
- Stanford, Craig; Allen, John S.; Antón, Susan C. (2009). Biological Anthropology: The Natural History of Humankind 2ª ed. Upper Saddle River, NJ: Pearson Prentice Hall. ISBN 978-0-13-601160-6. LCCN 2007052429. OCLC 187548835
- Strickberger, Monroe W. (2000). Evolution 3ª ed. Sudbury, MA: Jones and Bartlett Publishers. ISBN 978-0-7637-1066-8. LCCN 99032072. OCLC 41431683
- Stringer, Chris B. (1994). «Evolution of Early Humans». In: Jones, Steve; Martin, Robert D.; Pilbeam, David. The Cambridge Encyclopedia of Human Evolution 1ª paperback ed. Cambridge, UK: Cambridge University Press. ISBN 978-0-521-32370-3. LCCN 92018037. OCLC 444512451
- Swisher, Carl C. III; Curtis, Garniss H.; Lewin, Roger (2001). Java Man: How Two Geologists Changed Our Understanding of Human Evolution. Chicago: University of Chicago Press. ISBN 978-0-226-78734-3. LCCN 2001037337. OCLC 48066180
- Trent, Ronald J. (2005). Molecular Medicine: An Introductory Text 3ª ed. Burlington, MA: Elsevier Academic Press. ISBN 978-0-12-699057-7. LCCN 2004028087. OCLC 162577235
- Trevathan, Wenda R. (2011). Human Birth: An Evolutionary Perspective. New Brunswick, NJ: Transaction Publishers. ISBN 978-1-4128-1502-4. LCCN 2010038249. OCLC 669122326
- Ungar, Peter S.; Teaford, Mark F., eds. (2002). Human Diet: Its Origin and Evolution. Westport, CT: Bergin & Garvey. ISBN 978-0-89789-736-5. LCCN 2001043790. OCLC 537239907
- Walker, Alan (2007). «Early Hominin Diets: Overview and Historical Perspectives». In: Ungar, Peter. Evolution of the Human Diet: The Known, the Unknown, and the Unknowable. Col: Human Evolution Series. Oxford; New York: Oxford University Press. ISBN 978-0-19-518346-7. LCCN 2005036120. OCLC 132816551
- Wallace, David Rains (2004). Beasts of Eden: Walking Whales, Dawn Horses, and Other Enigmas of Mammal Evolution. Berkeley: University of California Press. ISBN 978-0-520-24684-3. LCCN 2003022857. OCLC 53254011
- Webster, Donovan (2010). Meeting the Family: One Man's Journey Through His Human Ancestry. Foreword by Spencer Wells. Washington, DC: National Geographic Society. ISBN 978-1-4262-0573-6. LCCN 2009050471. OCLC 429022321
- Wood, Bernard A. (2009). «Where Does the Genus Homo Begin, and How Would We Know?». In: Grine, Frederick E.; Fleagle, John G.; Leakey, Richard E. The First Humans – Origin and Early Evolution of the Genus Homo. Col: Vertebrate Paleobiology and Paleoanthropology. Dordrecht, the Netherlands: Springer Netherlands. pp. 17–28. ISBN 978-1-4020-9979-3. ISSN 1877-9077. LCCN 2009927083. OCLC 310400980. doi:10.1007/978-1-4020-9980-9_3 "Contributions from the Third Stony Brook Human Evolution Symposium and Workshop October 3–7, 2006."
Ligações externas[editar | editar código-fonte]
- O sistema imune humano pode limitar a evolução futura
- Insights sobre a evolução humana, a partir do genoma do gorila, 2012, em inglês
- Relações do Homo sapiens
- Espécies de Hominídeos em talkorigins.org
- DNA mostra que os Neandertais não são nossos Ancestrais
- «Neanderthais em Teste». Nova Online - Fornecido por PBS.
- «Tornando-se Humano». - Fornecido por PBS.
- Árvore da evolução de ancestrais fósseis do homem
- FOXP2 e a Evolução da Linguagem
- Atlas da Jornada Humana(National Geographic)
- Teoria sugere um papel maior ao melhor amigo do homem
- Novos fósseis de Jebel Irhoud, Marrocos e a origem pan-africana do Homo sapiens Jean-Jacques Hublin