肺
| 肺 | |
|---|---|
| ラテン語 | Pulmo |
| 英語 | Lung |
| 器官 |
循環器 呼吸器 |
| 動脈 | 肺動脈 |
| 静脈 | 肺静脈 |
| 神経 |
肺神経叢 胸肺枝 |
肺(はい、英: lung)は、脊椎動物の器官の1つである。肺臓とも呼ばれる。空気中から得た酸素を体内に取り込んだり、不要物である二酸化炭素を空気中に排出したりする役割(気泡の毛細血管で二酸化炭素の排出や酸素の吸収を行っている。)(呼吸)を持つ。これに対して水中から得た酸素を取り込み、水中に排出する器官が鰓(えら)である。
なお、脊椎動物以外の動物、いわゆる無脊椎動物においても、体内に一定の腔所を持ち、その内側でガス交換を行う器官をこう呼ぶ。節足動物のクモガタ類、軟体動物の腹足類にその例がある。
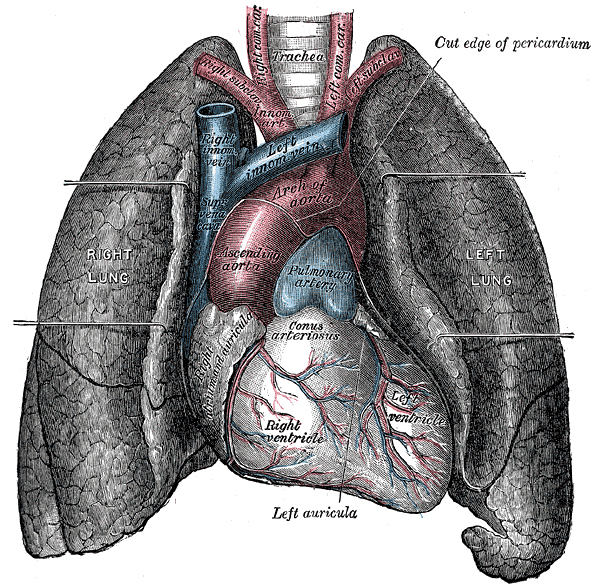
ヒト
[編集]ヒトの場合、重さは両肺で男性が約1kg、女性は約0.9kgほどあり、胸腔の大部分を占め[1]、主に気道と血管からなり、ガス交換のため両者は肺胞で接している。肺胞は約3億個で、総表面積は約70m2。
構造
[編集]横隔膜・肋間筋に囲まれた胸郭中にあり、肺の表面を覆っている漿膜を胸膜と言い[1]、横隔膜や肋間筋を裏打ちしている。肺を覆っている胸膜を臓側胸膜(肺胸膜、漿液である胸膜内液で満たされている[1]。
左と右、2つに分れているが、形状は左右対称にはなっておらず、右肺は上から順に上葉・中葉・下葉からなり、左肺はやや小さく上葉・下葉からなる[1]。この5つの肺葉を大葉と言い、大葉はさらに細かく10の肺区域に分けられる。それぞれ上部は尖っており、肺尖。下面は平らであり、肺底と呼ばれている。左肺に中葉がないのは、左右の肺を隔てる縦隔にある心臓が体幹の中心よりも左に寄っており、その分スペースが小さいためである。
気道と肺胞
[編集]口や鼻から入る空気の通り道を気道と言い[2]、咽頭で一つになり、喉頭で食道から前方に枝分れして気管となり[3]、縦隔で左右に枝分れして気管支になる[2]。
右肺は3葉あるので右気管支は気管から約25°の角度で枝分れする。左肺は心臓の分だけ上に寄っているので左気管支は気管から約35-45°の角度で枝分れする。 気管支は大葉へ向けて分岐し、さらに肺小葉に向けて分岐する。分枝を繰り返して軟骨を持たない複数弾性繊維を豊富に持つ終末細気管支を経て呼吸細気管支になり、その先端には肺胞がブドウのように密集している[1]。枝分かれは一定の法則に従って自己組織化するため、フラクタル構造になっている。
気管支には杯細胞や線毛細胞がある。細気管支にはクララ細胞がある。肺胞にはI型肺胞上皮細胞、II型肺胞上皮細胞がある。気管支にある杯細胞は気管粘液を出して湿度を保ち、線毛細胞は線毛運動によって吸気に混入した細菌等を咽頭へ流し戻す。これらの生理機能が正常に働いていれば肺胞は無菌に保たれているので、網細血管が直接空気と触れても細菌感染などは起こさない。
肺胞は、厚さ約0.1μmの扁平上皮である呼吸上皮細胞(肺胞上皮細胞)が直径約0.1-0.2mmの球状になり、空洞(肺胞気)を取り囲む構造を持つ。両肺合わせて約3億個がある肺胞では、内部に入り込んだ空気とそれを取り囲む多くの毛細血管の間でガス交換を行う場所であり、面積を広げると約70m2になる[1]。毛細血管は内皮細胞に走り、呼吸上皮細胞との間に基底膜がある。厚さ約0.5μmのこれら3層は血液空気関門と言う[1]。肺胞は弾性繊維で覆われ平滑筋を持たない。そのため、伸展はもっぱら気圧の変化によるものである[1]。肺胞でガス交換が行われる時は、I型肺胞上皮細胞が特に能動交換は行う訳ではなく、単にガス濃度の自然勾配によって受動交換が行われる。このため広い交換面積が必要になる。
肺胞上皮細胞内には、異物に対する免疫を持つマクロファージ(肺胞マクロファージ)や単球や、肺胞がひしゃげるのを防止するため脂質の表面活性物質を分泌するサーファクタント分泌細胞などがある[1]。肺胞にあるI型肺胞上皮細胞は薄い細胞で交換されるガスの通り道になっている。II型肺胞上皮細胞は厚い細胞で肺サーファクタントを出している。肺胞は極めて小さいので、そのままでは水の表面張力によって潰れてしまう。そのため表面活性物質を出して表面張力を下げて、肺胞が潰れない様にしている[1]。肺表面活性物質は胎生28週頃になってやっと出始めるため、妊娠28週以前に出産すると呼吸ができない新生児呼吸窮迫症候群 (RDS) になる危険性が極めて高い。そのような児のために現在は人工のサーファクタントを用いて、呼吸できる環境にするのが一般的である[1]。肺の血管内皮細胞はアンギオテンシン変換酵素 (ACE) を内分泌する。
大葉中ではお互いに穴でつながっているので、細菌性肺炎などを放置すると大葉性肺炎になる。
血管
[編集]流れる血管には大きく2系統があり、血液ガス交換のための系統を機能血管といい、いま1つは栄養血管と言う系統であり当臓器を養うためのものである[1]。
機能血管は心臓の右心室から肺動脈が出る。肺動脈は縦隔で左右に枝分れして右肺動脈と左肺動脈に分かれる。左右肺動脈は気道と同様に肺葉に向けて分岐して行き、最後は肺胞で毛細血管になる。肺胞でガス交換を終えた血管は分岐した時と同様に合流して行き、左右それぞれ2本の肺静脈となって左心房に流れ込む。栄養動脈である気管支循環系は、大動脈から直接分岐する。
生理
[編集]当臓器の膨張と収縮の仕組みは、横隔膜や肋間筋が胸腔を広げ、胸腔が陰圧になることで肺が立体的に引っ張られて受動的に膨らむ。一方縮むときは筋肉は使われず、当臓器の自ら縮もうとする力で収縮して空気の吸入・呼出をする。壁側胸膜は知覚神経が豊富で、肺が痛む時はこの神経が関与している。
なお、サーファクタント(界面活性剤)が、肺の円滑な動きには必要である。このサーファクタントが不足した状態になったことが新生児呼吸窮迫症候群の原因である。サーファクタントが足りないために上手く肺胞が広がらず、ガス交換が上手くゆかなくなっているのである。なお新生児呼吸窮迫症候群は、妊娠期間が短いほど(早産であるほど)発生しやすく、新生児が死亡する原因の1つとなる。
CT解剖学
[編集]気管支と肺動脈は原則として隣接し平行に走行する。肺区域、亜区域、小葉の中心を走行する。これに対して肺静脈はこれらの境界を走行する。CTでは気管支に隣接する血管が肺動脈であり、肺動脈と肺動脈の間にある血管が肺静脈である。正常なヒトでは気管支は亜区域までしか追うことはできないのでそこまでは有効な方法である。機能動脈は肺動脈だが、それ以外に栄養血管として気管支動脈が存在する。気管支動脈は下行大動脈から直接分枝するが正常では細いため造影CTでその近位部が確認されるにすぎない。
当臓器はリンパが豊富な組織である。気管支周囲、肺血管周囲、小葉間隔壁、胸膜の間質に分布している。特によく発達しているのが、気管支周囲と肺動脈周囲である。基本的には肺末梢から肺門部に向かって流れている。リンパ管そのものはCTでは確認できないが、癌性リンパ管炎や鬱血性心不全のようにリンパ浮腫を起こすと、気管支壁が肥厚し、血管陰影が拡大し小葉間隔壁が確認できるようになる。
構造を理解する上で欠かせない概念が二次小葉といわれるものである。もっとも有名なものはMillerによる定義である。二次小葉の中央を気管支と肺動脈が小葉間隔壁の中を肺静脈が走っている。肉眼的にも確認ができる小葉間隔壁に囲まれた多面体である。この概念は間質性病変を理解するのに役に立つ。二次小葉は30個ほどの細葉が集まってできているとされている細葉はCTでは確認ができない。
亜区域を同定するには気管支を辿っていくのがわかりやすい。原則として区域気管支の番号と肺区域の番号は一致し、大体気管支が肺区域の中央を通過することを念頭におくと手術後や偏位のある場合でも亜区域を同定できる。
- 右上葉
- 右主気管支はまず、上方に右上葉気管支を分枝する、右上葉気管支は上方(外側)にB1、後方にB2、前方にB3の分枝をする。反時計回りに番号が振られていることに注意が必要である。B1はS1(肺尖区)の区域気管支であり、B2はS2(後上葉区)、B3はS3(前上葉区)の区域気管支である。右主気管支は右上葉気管支を分枝した後、中間気管支管(左には存在しない)という。
- 右中葉
- 中間気管支管は前方に右中葉枝を分枝する。右中葉枝は外側(前方)のB4と内側(後方)のB5に分枝する。B4はS4(外側中葉区)、B5はS5(内側中葉区)の区域気管支である。
- 右下葉
- 右中葉枝を分枝した直後、後方にB6が分枝される。次いで、B7が前下内方へ、B8が前下外方へB9+10が下後方に分枝する。B6はS6(上下葉区)、B7はS7(内側肺底区)、B8はS8(前肺底区)、B9+10はS9(外側肺底区)、S10(後肺底区)の区域気管支である。
- 左上葉
- 左主気管支から左上葉気管支が上方に分枝する。左上葉気管支は上行する上区枝と前下方の舌区枝に分枝する。上区枝は上後方のB1+2と前方のB3に分枝する。舌区枝は前方のB4と下外方のB5に分かれる。B1+2はS1+2(肺尖後区)、B3はS3(前上葉区)、B4はS4(上舌区)、B5はS5(下舌区)の区域気管支である。
- 左下葉
- 左上葉気管支を分枝した直後、B6を後方に分枝する。左側にはS7(内側肺底区)は存在しないことが多く、前下方のB8、後方のB9+10を分枝する。B9+10は外側のB9と内側のB10に分枝する。B6はS6(上下葉区)、B8はS8(前肺底区)、B9はS9(外側肺底区)、B10はS10(後肺底区)の区域気管支である。
脊椎動物
[編集]起源
[編集]酸素の摂取は水中からより空気中からのほうがはるかに有利となる。これは水中における溶存酸素がせいぜい15℃で7ml/lであるのに対して空気中には209ml/lにも達すること、空気の比重は水の1000分の1、空気の粘性は水の100分の1であること、酸素の血液中への拡散速度は空気中が水中の50万倍にも上ることによる。しかし、初期脊椎動物によって水呼吸に用いられてきた鰓は水との接触面積を柔軟で微細な襞状構造によって達成しているため、これを空気中に引き出すと襞間の水の凝縮力によってしぼんでしまい、呼吸媒体との接触面積が水中に比して著しく低下する。そのため、空気呼吸の利点を享受するには新たな呼吸器官が必要となる。この空気呼吸器官は鰓のような凸状の酸素摂取面を体外に発達させるのではなく、体内に凹状の酸素摂取面を設けることで、空気の圧力により呼吸媒体との接触面積が広く維持できる。こうして鰓裂直後の消化管腹壁より分化した嚢状器官として肺は発生した。脊椎動物が肺を獲得した時期に関しては初期の有顎動物下門である板皮綱に既に肺を持つものがいたとする説があるが、初期の硬骨魚綱には確実に存在している。また、軟骨魚綱は肺を持つことがなかったグループである。
肺循環の分化
[編集]発達と同時に、取り入れた酸素を機能的に全身に運ぶため、血管の配置も変化している。魚類では心臓からでた動脈の一つが通り、その後全身に巡るという形を取るが、両生類以上では心臓から来て当臓器を通って、すぐに心臓へ戻り、あらためて全身へ運ばれるという形を取る。つまり心臓から全身を回る循環と、肺へ行ってすぐ戻る循環が分化する。これを、それぞれ体循環と肺循環という。ただし、これを十分に行うためには心臓内部が二つに分かれなければならないが、そのための壁は爬虫類以上でなければ完成されない。
| 種 | 温度 (℃) |
O2摂取(%) | CO2排出(%) | ||
|---|---|---|---|---|---|
| 空気から | 水から | 空気へ | 水へ | ||
| 肺魚類 | |||||
| プロトプテルス・エチオピクス | 24 | 92 | 8 | 32 | 68 |
| レピドシレン・パラドクサ | 20 | 42 | 58 | 0 | 100 |
| ガーパイク類 | |||||
| ロングノーズガー | 22 | 73 | 27 | 8 | 92 |
| スポッテッドガー | 20 | 42 | 58 | 0 | 100 |
| アミア類 | |||||
| アミア | 30 | 74 | 26 | 39 | 61 |
| 20 | 69 | 31 | 23 | 77 | |
| 10 | 8 | 92 | 7 | 93 | |
| 30 | 40 | 60 | 8 | 92 | |
| オステオグロッスム類 | |||||
| ピラルクー | 28~30 | 78 | 22 | 37 | 63 |
両生類
[編集]基本的に魚類と同じ単純な袋状の構造で、通常は最大の呼吸器官であるが、これだけでは全呼吸量をまかなうことができない。両生類の皮膚の角質層はきわめて薄く、また通常粘液で覆われているので酸素や二酸化炭素が透過しやすく皮膚呼吸の能力も高い。皮膚呼吸への依存度は活動時のカエルで全呼吸量の2分の1から3分の1程度と言われている。
水生種の中には幼形成熟(ネオテニー)により
両生類の中では活動的で呼吸量も多いカエルのものは、比較的複雑になっており、内壁の襞の発達によりいくつもの泡沫が集合したような外観である。
爬虫類
[編集]爬虫類は基本的にほぼ当臓器に依存した呼吸が可能な程度に構造が複雑化して、内壁は海綿状にまで発達している。中には哺乳類と同程度にまで複雑化したものを持つものもある。皮膚の表面には厚い角質が発達するので皮膚呼吸への依存は低いが、水生のカメなどでは咽頭や総排泄孔の内壁の粘膜を水中での補助呼吸器官として用いている。さらにワニやカメなどが潜水するときは心臓の操作によって肺循環を低下させ、肺呼吸を停止した状態での効果的な血液循環を図ることが知られている。また、ヘビの場合、著しく細長くなった体制に応じて左の肺が退化し、右の肺のみが発達する。
鳥類
[編集]鳥類は、気嚢と肺管の組み合わせによる極めて効率的なガス交換システムをもつ。
気管から分岐した気管支は、最終的に並行して走る肺管と呼ばれる細かい管になり、これがガス交換の場となる。ここから5対の気嚢と呼ばれる大きな袋が出ている。
呼吸運動において5対の気嚢がふいごのように伸縮するが、肺管は伸縮しない。吸気はまず3対の後気嚢に直接吸い込まれ、保持される。次に酸素を豊富に含んだ後気嚢内の空気が後方から肺管に送り込まれる。肺管の血管には前方から血流が送られており、肺管の前方の古い空気との間で二酸化炭素を排出した後、後方の新しい空気から酸素を吸収する。肺管を通過した空気は前気嚢に集められ、気管に向けて排出される。
このようにガス交換部における空気の流れは一方向のみで、常に新鮮な空気が血液と対向方向に流れる。他の脊椎動物では空気は往復運動している。そのためガス交換部である肺胞に常にある程度の古い空気が残存せざるを得ず、新鮮な空気は常にかなりの量の古い空気と混合した後にガス交換が行われることになる。
こうした効果的なガス交換能力を身につけていたことが鳥の激しいエネルギー消費を伴う飛行を可能にしたと言える。しかし、鳥の気嚢は全身にくまなく入り込んでいるため、獣医学的には鳥の呼吸器感染症は重篤になりやすいと言われている。
恐竜
[編集]鳥類は獣脚類の恐竜から分岐して進化した。現存鳥類の呼吸システムを、獣脚類(もしくは恐竜全体)が既に持っていたという仮説があり、研究がすすめられている。2005年には、マジュンガサウルス脊椎骨の構造の研究から獣脚類が気嚢を持つ証拠が提出され、この仮説の実証が前進した。
哺乳類
[編集]鳥類以外の動物の当臓器の基本型ともいえ、哺乳類のものは最も複雑な構造を持つ。気管からは細気管支が幾度も分枝を重ね、膨大な数の微細な肺胞に至り、ここがガス交換の場となる。著しいガス交換面積の獲得により酸素の摂取能力と二酸化炭素の排出能力は非常に高くなっている。ヒトも哺乳類の一種であるため、基本的な構造と機構については上記のヒトの項目を参照のこと。
無脊椎動物
[編集]脊椎動物以外の動物にも「肺」の名が付く呼吸器はあるが、いずれも上述の肺とは別起源の器官である。
- 節足動物クモガタ類の場合
- 蛛肺類に分類されるクモガタ類は、書肺(しょはい)という対になる呼吸器を後体の腹面に配置される[5]。その中でサソリは4対、ウデムシ・サソリモドキ・クモ下目以外のクモは2対、ヤイトムシとクモ下目のクモは1対をもつ[5]。クモ下目の中では、ユウレイグモ科など一部を除いて昆虫と相似の気管系を書肺と併用している。体サイズの小さな節足動物では、肺のような血液循環に依存した呼吸器よりも、気管系のような体組織に直接酸素を届ける呼吸器のほうが効率がよいことが知られている。書肺は、クモガタ類と同じく鋏角類の節足動物であるカブトガニ類に見られるような書鰓(しょさい)に起源する器官と考えられる[6][7]。いずれも関節肢由来の蓋板の内側に配置された、本のページのように畳んだ無数の薄板によって構成される[6][7]。ただし能動的で水中呼吸用の書鰓とは異なり、空気呼吸用の書肺は体の内側に占め込んで不可動になり、各薄板の間には、空気を取り込める隙間を支えるための小柱がたくさん生えている[6][7][8]。
- 軟体動物腹足類の場合
- カタツムリなど、陸生種を含む有肺類では、殻の入り口近くの外套膜の下に、空気の出入りできる腔所があり、ここで空気呼吸する。これを肺という。通常の腹足類では鰓を収納している外套腔に空気を吸い込むようになったものである。空気の出入り口は、殻の口のそば、体の側面に1つだけある。有肺類にはモノアラガイのような淡水生種やカラマツガイのような海生種も知られているが、前者は皮膚呼吸のほか時々水面上に呼吸孔を開いて肺呼吸をし、後者は二次的な鰓をもち海水から酸素を取り込む。有肺類以外の腹足類でもヤマキサゴ、ヤマタニシのような陸生種や、スクミリンゴガイのような貧酸素になりやすい淡水に生息する種にも空気呼吸するものがあり、これらも外套腔に空気を吸い込んでガス交換を行えるようになっている。
脚注
[編集]- ^ a b c d e f g h i j k l 佐藤・佐伯(2009)、p.87-47、第4章 呼吸、2.呼吸器系の構造と機能、5)肺lung
- ^ a b 佐藤・佐伯(2009)、p.86-87、第4章 呼吸、2.呼吸器系の構造と機能、4)気管tracheaと気管支bronchus
- ^ 佐藤・佐伯(2009)、p.83-84、第4章 呼吸、2.呼吸器系の構造と機能、2)咽頭pharynxと喉頭larynx
- ^ 参考文献:板沢靖男・羽生功 / 編『魚類生理学』(恒星社厚生閣、1991年(平成3年))
ISBN 4-7699-0706-0
P.31の表よりデータを抜粋編集 - ^ a b Dunlop, Jason A.; Lamsdell, James C. (2017). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3): 395. ISSN 1467-8039.
- ^ a b c Scholtz, Gerhard; Kamenz, Carsten (2006). “The book lungs of Scorpiones and Tetrapulmonata (Chelicerata, Arachnida): evidence for homology and a single terrestrialisation event of a common arachnid ancestor”. Zoology (Jena, Germany) 109 (1): 2–13. doi:10.1016/j.zool.2005.06.003. ISSN 0944-2006. PMID 16386884.
- ^ a b c Lamsdell, James C.; McCoy, Victoria E.; Perron-Feller, Opal A.; Hopkins, Melanie J. (2020-11-02). “Air Breathing in an Exceptionally Preserved 340-Million-Year-Old Sea Scorpion” (English). Current Biology 30 (21): 4316–4321.e2. doi:10.1016/j.cub.2020.08.034. ISSN 0960-9822. PMID 32916114.
- ^ Kamenz, Carsten; Dunlop, Jason A; Scholtz, Gerhard; Kerp, Hans; Hass, Hagen (2008-04-23). “Microanatomy of Early Devonian book lungs” (英語). Biology Letters 4 (2): 212–215. doi:10.1098/rsbl.2007.0597. ISSN 1744-9561. PMC 2429929. PMID 18198139.
参考文献
[編集]- 監修:佐藤昭夫、佐伯由香『人体の構造と機能 第2版』(第2版第6刷)医歯薬出版、2009年。ISBN 978-4-263-23434-1。
外部リンク
[編集]- 呼吸器系の構造と機能 (ビジュアル生理学 内の項目)
呼吸器系の正常構造・生理 | |||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 気道系 |
| ||||||||||||||||||||||||||||||
| 血管系 |
| ||||||||||||||||||||||||||||||
| 運動器系 |
| ||||||||||||||||||||||||||||||
| 神経系 |
| ||||||||||||||||||||||||||||||
|
| |||||||||||||||||||||||||||||||

| 国立図書館 | |
|---|---|
| その他 | |